 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 103, No. 2, March, 2008 , pp. 216-219 Short Communication Reproductive isolation between sympatric and allopatric Brazilian populations of Lutzomyia longipalpis s.l. (Diptera: Psychodidae) Nataly A Souza, Claudia A Andrade-Coelho, Felipe M Vigoder1, Richard D Ward2, Alexandre A Peixoto1/+ Laboratório
de Transmissores de Leishmanioses 1Laboratório
de Biologia Molecular de Insetos, Instituto Oswaldo Cruz-Fiocruz,
Av. Brasil 4365, 21045-900 Rio de Janeiro, Brasil 2Institute
of Science and Technology in Medicine, School of Life Sciences,
Keele University, Staffordshire, U.K. Financial support: CNPq, Fiocruz (PAPES IV), Howard Hughes Medical Institute Received
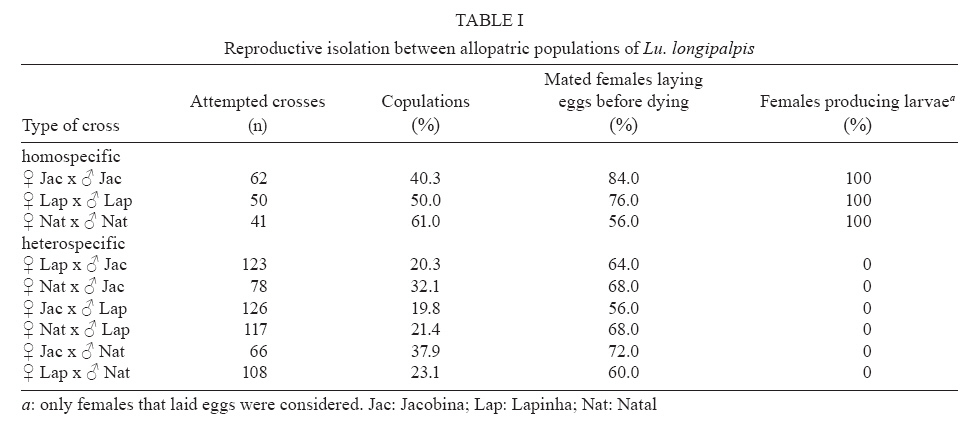
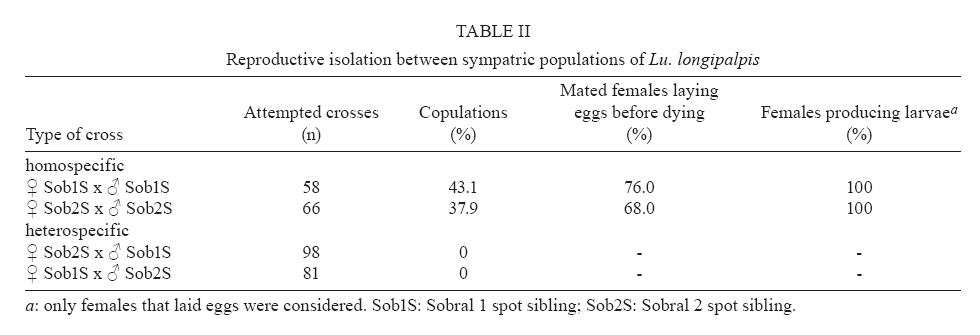
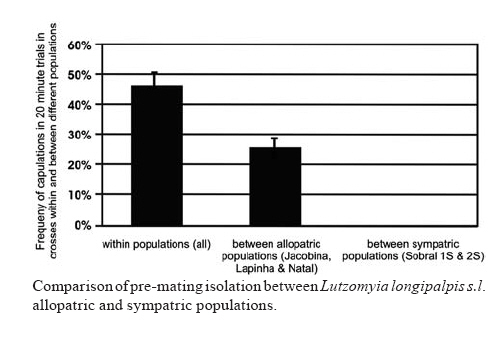
21 September 2007 Code Number: oc08038 Lutzomyia longipalpis s.l., the main vector of Leishmania chagasi in Latin America, is a species complex although the exact number of siblings is yet unknown. In Brazil, the siblings differ in male copulatory courtship songs and pheromones that most certainly act as pre-zygotic reproductive barriers. Here we analysed the reproductive isolation between three allopatric and two sympatric populations of Lu. longipalpis s.l. from Brazil. The results indicate a strong copulatory and pre-mating isolation between the three allopatric populations. In addition, the results also indicate a stronger pre-mating isolation between the two sympatric siblings than between the three allopatric ones, suggesting a role for reinforcement in the speciation of the Lu. longipalpis s.l. complex. Key words: visceral leishmaniasis - sand flies - species complex - reinforcement Lutzomyia longipalpis s.l. occurs from Argentina to Mexico (Young & Duncan 1994) and because of its wide distribution there have been doubts expressed that this taxon is a single species. Mangabeira (1969) commented on morphological variation seen in males. He observed that specimens captured in the state of Pará, North Brazil, bore a single pair of pale tergal spots on abdominal segment IV (1S males) and those captured from the Northeastern state of Ceará (CE) showed an additional pair of spots on segment III (2S males). Mangabeira commented that the two forms were found in different ecological conditions and speculated that they might represent two species. The existence of a complex was confirmed by Ward et al. (1983, 1988) and Lanzaro et al. (1993) in crossing experiments, although the male spot phenotype turned out to be only useful as a morphological marker of different species in some localities such as Sobral, CE (see below) (reviewed in Bauzer et al. 2007). In Brazil, Lutzomyia longipalpis s. l. (Lutz & Neiva, 1912) is without any doubt the principal vector of American visceral leishmaniasis (AVL). It is present in nearly all of the foci of visceral leishmaniasis, is highly anthropophilic as well as readily feeding on dogs and foxes which act as reservoirs of Leishamania chagasi which are frequently found infected (reviewed in Lainson & Rangel 2005). Analysis of male copulatory courtship songs (Souza et al. 2002, 2004) and pheromones (Ward et al. 1988, Hamilton et al. 1996a,b, 2005) have shown that different Brazilian populations of Lu. longipalpis belong to different species. Their sibling species status was also confirmed by the use of molecular markers, such as the behavioural genes period (Bauzer et al. 2002a,b) and cacophony (Bottecchia et al. 2004), and microsatellites (Maingon et al. 2003, Watts et al. 2005). Therefore, there is now no doubt that Lu. longipalpis s.l. is a species complex in Brazil (reviewed in Bauzer et al. 2007, Maingon et al. 2008), a fact that might have significant epidemiological consequences as sibling species often differ in their importance as vectors. The best studied populations in Brazil, from an evolutionary point of view, are located in Jacobina (state of Bahía), Lapinha Cave in Lagoa Santa (state of Minas Gerais), Natal (state of Rio Grande do Norte) and Sobral (CE) (Bauzer et al. 2007). The first three localities represent different allopatric siblings (Bauzer et al. 2002b, Bottecchia et al. 2004, Watts et al. 2005) while in Sobral two species coexist in sympatry (Bauzer et al. 2002a, Maingon et al. 2003, Bottecchia et al. 2004). In the present study we compared the reproductive isolation among the three allopatric populations and between the Sobral sympatric siblings. Initially samples of the populations of Jacobina, Lapinha and Natal were obtained using CDC traps (4 to 9 traps in each locality). All males collected in Lapinha had a single pair of spots (1S) in segment IV and all males from Jacobina were 2S (two pairs in segments III and IV, with similar sizes). The majority of males from Natal had the intermediate phenotype, with the extra pair in segment III with less than 3/4 of the size of the spots in segment IV (see Ward et al. 1988). The population from Natal is highly polymorphic for the male spot phenotype but in this locality there is no evidence for the existence of sympatric sibling species (Mukhopadhyay et al. 1998, Bauzer et al. 2002b, Bottecchia et al. 2004, Watts et al. 2005). Eggs from 100 to 200 wild-caught females from each of the three localities were used to establish laboratory colonies following the methodologies of Rangel et al. (1985) and Souza et al. (1999). Crossing experiments (see below) were carried out using mainly sand flies from the F2 and F3 generations. After these initial crosses three new samples from each locality were obtained to boost the colonies and new crossing experiments were carried out after each new collection. The establishment of colonies of the two sympatric populations of Sobral occurred more than one year after the end of the crossing experiments with allopatric populations. By this time the three original allopatric colonies were no longer available. Sobral is one of the few localities were the number of male tergal spots (1S or 2S) can be used to separate sympatric siblings (Ward et al. 1988, Bauzer et al. 2002a, Maingon et al. 2003). Sand flies were collected using 9 CDC traps in three different houses, distant approximately 3-11 km from each other. Initially more than 170 isofemale lines (IL) were established from wild caught females. These IL were sorted according to the number of spots in their F1 males. A few lines (~ 4%) exhibiting ambiguous intermediate phenotypes (Ward et al. 1988) were observed and discarded before colony establishment. 1S and 2S colonies were established and maintained from the sorted IL (~ 34% 1S and ~ 62% 2S) as described in Rangel et al. (1985) and Souza et al. (1999). The initial crosses were carried out using mainly the F2 and F3 generations of these two colonies. As before, three new samples were obtained from Sobral to boost the colonies and new crosses were carried out after each new collection. For the crossing experiments pupae were separated in individual vials before emergence so males and females were virgin when they were used in the crosses. Sand flies 5-7-day old were used in the experiments. One male and one blood-fed female were placed together in a small mating chamber (15 mm x 10 mm x 5 mm) for up to 20 min. If no copulation occurred in 20 min the couple was discarded and replaced until a total of 25 copulations were observed for each cross. Heterospecific and homospecific crosses were usually carried out in parallel. The females that copulated were individualized again in oviposition vials and post blood-feeding egg hatching was monitored. Table I shows the number of attempted crosses, the percentage of copulations observed, the percentage of females that laid eggs before dying and the percentage of those that produced larvae in crosses between the three allopatric populations. The results show that all females that were mated to homospecific males and survived long enough to lay eggs produced viable larvae. On the other hand none of the heterospecific crosses yield progeny. Table II shows similar results for crossing experiments carried out with the sympatric siblings of Sobral. As before, all females that were mated to homospecific males, and that did not die before laying eggs, successfully produced larvae. Similar results were obtained by Ward et al. (1983, 1988) who observed insemination failure in copulations between males and females of some Brazilian populations, including the Sobral siblings. Interestingly enough, Lu. Longipalpis s.l. males produce copulatory "lovesongs" (Souza et al. 2002, 2004) and pheromones (Hamilton et al. 1996a,b, 2005, Casanova et al. 2006) that show striking differences between populations suggesting that those acoustic and chemical signals have a role in the observed insemination failures. The results presented in Tables I, and II also show evidence for pre-mating reproductive isolation as summarized in the Figure. While copulations were observed in nearly half of the attempted homospecific crosses (0.465 ± 0.042), only about 25% (0.258 ± 0.030) of copulations were observed in heterospecific crosses among allopatric populations. This difference is highly significant (t = 4.125; d.f. = 9; p < 0.01; after arcsine transformation) and indicates pre-mating reproductive isolation. Moreover, the results obtained with the heterospecific crosses involving the two sympatric populations of Sobral indicate an even stronger pre-mating isolation between the two siblings in this locality as no copulations were observed in a total of 179 crosses. The difference between sympatric and allopatric heterospecific crosses is also highly significant (t = 8.56; d.f. = 6; p < 0.001; after arcsine transformation). Analysis of the divergence and gene flow among Brazilian populations using a number of different markers suggest a recent speciation process with the possible occurrence of introgression events (Bauzer et al. 2002a,b, Maingon et al. 2003, Bottecchia et al. 2004, Watts et al. 2005). The fact that sympatric siblings show stronger pre-mating isolation than the allopatric populations suggests that reinforcement of reproductive isolation (reviewed in Servedio & Noor 2003) might have had a role in the speciation of Lu. longipalpis complex siblings in Brazil. We are currently expanding our population genetic analysis of this complex to other populations. It will be interesting to extend these crossing experiments to other areas where sympatric siblings might be found to determine how widespread the occurrence of enhanced pre-mating isolation is in these populations when compared to allopatric siblings. ACKNOWLEDGMENTS To Maria de Lurdes dos Santos (IOC-Fiocruz) for technical assistance in the Insectary, and Luiz Henrique Costa (IOC-Fiocruz), Lindenberg de Souza (Secretaria de Saúde do Estado do Ceará), Iraci Duarte (Secretaria Estadual de Saúde Pública do Rio Grande do Norte), Arlinete Medeiros (Secretaria Estadual de Saúde Pública do Rio Grande do Norte), Ítalo Sherlock (CPqGM-Fiocruz) and Reginaldo Brazil (IOC-Fiocruz) for assistance with the sand fly field collections. REFERENCES
Copyright 2008 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc08038t1.jpg] [oc08038t2.jpg] [oc08038f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}