 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
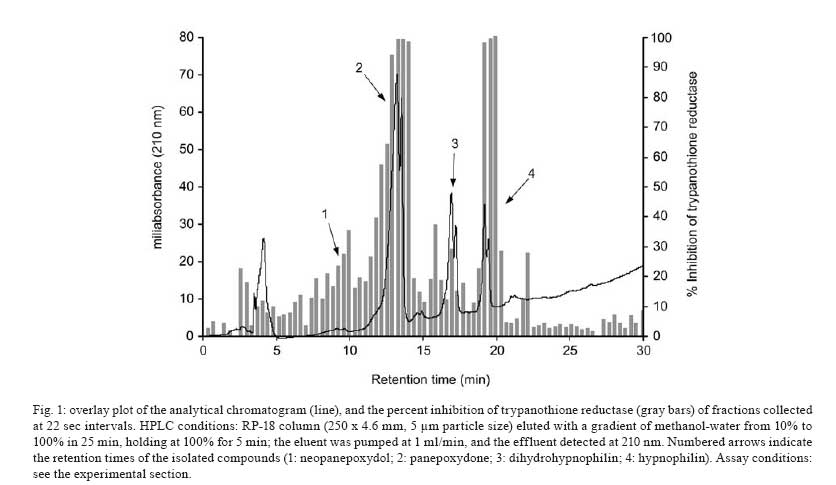
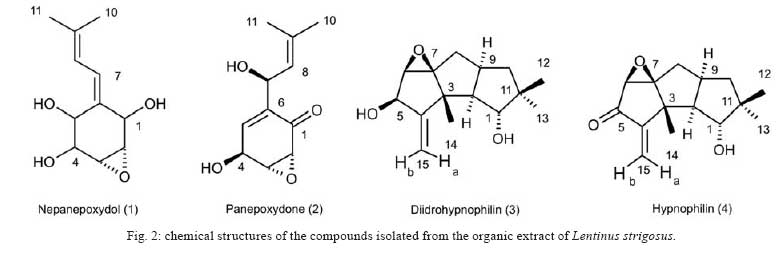
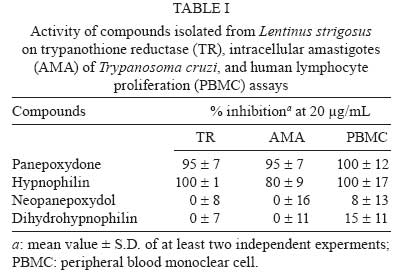
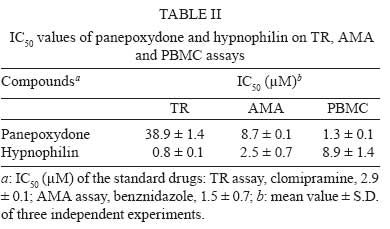
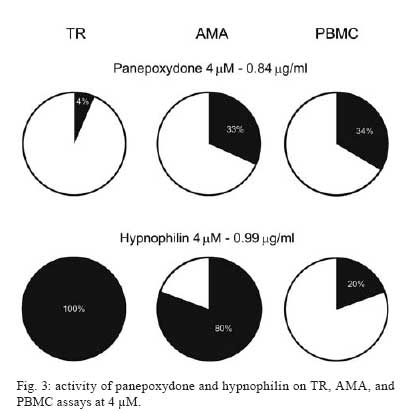
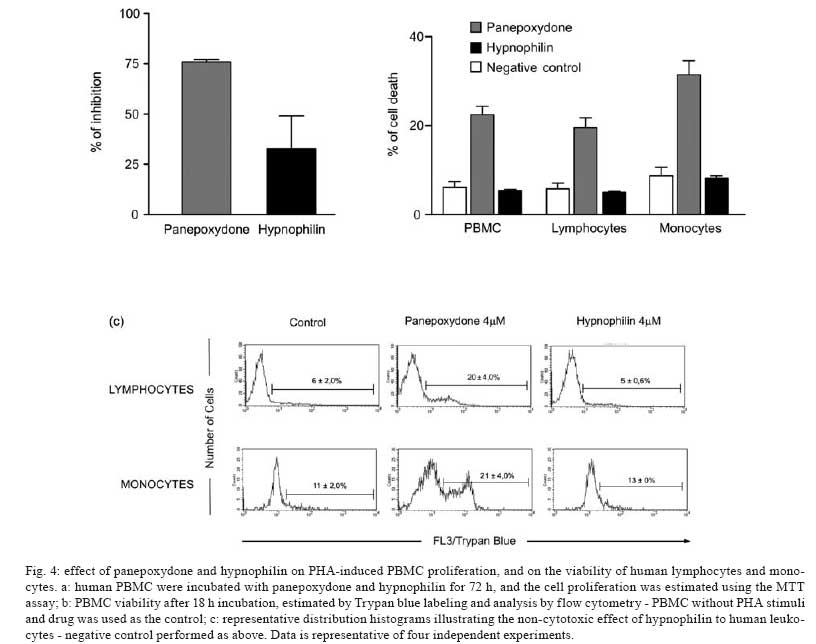
Memórias do Instituto Oswaldo Cruz, Vol. 103, No. 3, May 2008, pp. 263-270 A potent trypanocidal component from the fungus Lentinus strigosus inhibits trypanothione reductase and modulates PBMC proliferation Betania Barros CotaI; Luiz Henrique RosaI, ++; Elaine Maria Souza FagundesI, +++; Olindo Assis Martins-FilhoII; Rodrigo Correa-OliveiraIII; Alvaro José RomanhaIV; Carlos Augusto RosaV; Carlos Leomar ZaniI, + ILaboratório de Química de Produtos Naturais Financial support: Fapemig, Capes, CNPq, Fiocruz Received 12 December 2007 Code Number: oc08045 ABSTRACT The fungus Lentinus strigosus (Pegler 1983) (Polyporaceae, basidiomycete) was selected in a screen for inhibitory activity on Trypanosoma cruzi trypanothione reductase (TR). The crude extract of L. strigosus was able to completely inhibit TR at 20 µg/ml. Two triquinane sesquiterpenoids (dihydrohypnophilin and hypnophilin), in addition to two panepoxydol derivatives (neopanepoxydol and panepoxydone), were isolated using a bioassay-guided fractionation protocol. Hypnophilin and panepoxydone displayed IC50 values of 0.8 and 38.9 µM in the TR assay, respectively, while the other two compounds were inactive. The activity of hypnophilin was confirmed in a secondary assay with the intracellular amastigote forms of T. cruzi, in which it presented an IC50 value of 2.5 µ M. Quantitative flow cytometry experiments demonstrated that hypnophilin at 4 µM also reduced the proliferation of human peripheral blood monocluear cells (PBMC) stimulated with phytohemaglutinin, without any apparent interference on the viability of lymphocytes and monocytes. As the host immune response plays a pivotal role in the adverse events triggered by antigen release during treatment with trypanocidal drugs, the ability of hypnophilin to kill the intracellular forms of T. cruzi while modulating human PBMC proliferation suggests that this terpenoid may be a promising prototype for the development of new chemotherapeutical agents for Chagas disease. Key words: fungal natural products - Chagas disease - drug discovery - immunomodulators - Basidiomycota Chagas disease is caused by the protozoan parasite Trypanosoma cruzi and affects millions of people in Latin America, having an enormous economic and social impact in the endemic areas. These patients rely on treatment with nitrofuran (nifurtimox; Bayer) or nitroimidazol (benznidazole; Roche), medicines that display little or no activity in chronic infections, in spite of their beneficial effect during the acute phase of the disease (Cançado 2002, Coura & Castro 2002). However, significant differences in the therapeutic effectiveness are observed between these two drugs, especially when considering distinct geographical areas, which is probably due to the occurrence of naturally resistant and susceptible T. cruzi strains (Filardi & Brener 1987, Toledo et al. 2003). Furthermore, both drugs cause several side effects that contribute to their reduced use in clinical medicine (Cançado 1985). Thus, new compounds are needed to develop more effective medicines to treat Chagas disease, especially in its chronic phase, is needed (Schmidt & Krauth-Siegel 2002, Nwaka & Ridley 2003). However, due to low profit prospects, the development of new trypanocidal drugs is not attractive to the pharmaceutical industry (Nwaka & Ridley 2003). Thus, this endeavor is being conducted mainly in academic laboratories (Fairlamb 1999). Among the many different strategies for drug discovery, screening the local biodiversity for bioactive natural products using appropriate bioassays is an interesting alternative for researchers in affected areas, especially in countries with a rich biodiversity. There is a vast literature describing that natural products produced by plants, fungi and other organisms are the direct source of or inspiration for many of the currently available medicines (Newman & Cragg 2007). In this regard, our group screens the Brazilian biodiversity for new bioactive natural products using in vitro bioassays related to neglected diseases, especially Chagas disease and leishmaniasis (Ribeiro et al. 1997, Rosa et al. 2005, 2006). We evaluated more than 3,000 extracts from plant and fungi using an inhibition assay with trypanothione reductase (TR) (unpublished results). This enzyme is a validated molecular target in Trypanosoma and Leishmania species (Fairlamb et al. 1985, Dumas et al. 1997, Tovar et al. 1998, Krieger et al. 2000). As a result of the screening, we found that the ethyl acetate extract of the culture of Lentinus strigosus, a basidiomycete fungus belonging to the Polyporaceae family (Pegler 1983), was able to inhibit 100% of the enzyme activity at 20 µg/ml. Species from the genus Lentinus are normally saprophytic, lamellate wood-decaying basidiomycetes (Kirk et al. 2001). Most of its 40 species grow in the tropics, although some can be found in the boreal region (Pegler 1983). Previous investigations on the chemical composition of Lentinus species identified several sesquiterpenes from L. lepideus (Hanssen 1982, 1985) and L. connatus (Rukachaisirikul et al. 2005) and benzoquinones from L. degener (Anchel et al. 1948) and L. adhaerens (Lauer et al. 1991). However, to the best of our knowledge, no investigations studying TR inhibitors were performed with any species from this genus, and this is the first study on the species L. strigosus. Here, we describe the bioassay-guided isolation and identification of the active compounds present in L. strigosus extract. We also show that one of these compounds, the terpenoid hypnophilin, has an interesting biological activity profile, making it an attractive lead for further investigations of drugs to treat Chagas disease. MATERIAL AND METHODS General experimental procedures - The optical rotations were measured on a Perkin-Elmer 341 polarimeter (25ºC, Na lamp, 589 nm). Infrared spectra were obtained on a Shimadzu FTIR-8400, with the samples in KBr pellets. 1H (400 MHz), 13CNMR (100 MHz), DEPT, HMQC, and HMBC NMR spectra were measured on a Brucker DRX 400 spectrometer. Electron impact (70 eV) low-resolution mass spectra (EI-MS) were measured with a Shimadzu QP5050A spectrometer, equipped with a direct insertion probe. Electrospray ionization mass spectra (ESI-MS) were measured on a Thermo Finnigan LCQ-Advantage spectrometer. Biological material - The L. strigosus (Schwein) Fr. isolate was deposited in the UFMGCB collection (Universidade Federal de Minas Gerais, Brazil) and received the code UFMGCB975. The specimen was identified using classical taxonomic methods (Pegler 1983). Fungus fermentation - The fungus was grown on malt extract agar (MEA, Difco, US) and transferred (three 5 mm discs) to ten 250 ml Erlenmeyer flasks containing 25 ml of MEC medium (malt extract 2%, peptone 0.1%, glucose 1.5%). The flasks were shaken at 150 rpm and 28ºC for five days. The contents of these culture flasks were then transferred to an 8 l bioreactor and cultivated for nine days at 28ºC. Extract preparation - The mycelium was separated from the culture by vacuum filtration, and the filtrate was extracted with ethyl acetate (5 x 300 ml for each 1,000 ml of broth). The organic phase was separated and dried over anhydrous sodium sulfate. After filtration, the solution was concentrated in a rotary evaporator at temperatures below 45ºC. The extract was then dried in a vacuum centrifuge at 40ºC to yield 2.29 g of a brown gum. An aliquot was used to prepare a stock solution at 20 mg/ml in dimethylsulphoxide (DMSO), which was used in all biological assays. Both the crude extract and stock solution were stored at -40ºC. Isolation and purification - Initially, 100 µg of the crude extract was separated in an analytical HPLC column (Shim-pack ODS, 5 µm, 4.6 x 250 mm), using a linear gradient from 10 to 100% CH3OH in 25 min at 1.0 ml/min. The effluent was monitored with a photodiode array UV detector and collected in a 96-well plate (220 µl per well, 80 wells). The plates were dried in an oven at 40ºC and then subjected to the TR bioassay to identify the fractions containing metabolites with inhibitory activity (Fig. 1). A semi-preparative fractionation was then performed in a 20 x 250 mm reverse-phase HPLC column (Shim-pack prep-ODS, 5 µm), using a 34 min linear gradient from 15 to 100% CH3CN in H2O. Five aliquots of approximately 100 mg of crude extract were fractionated at a flow rate of 10 ml/min, with UV detection at 210 and 254 nm. One fraction afforded a white amorphous solid that, upon crystallization with a mixture of CH2Cl2-Hex, yielded 32 mg of dihydrohypnophilin (3). The remaining fractions were further fractionated by semi-preparative isocratic normal-phase HPLC (Shim-pack prep-Si, 5 µm, column 20 x 250mm), eluted with CH2Cl2-MeOH (95:5 or 97:3) at 10 ml/min. This procedure yielded 70 mg of panepoxydone (2), 3.8 mg of neopanepoxydol (1), and 14 mg of hypnophilin (4). Neopanepoxydol (1) - White amorphous powder; IR (KBr): vmax = 3308 (OH), 2962 (aliph.), 2862 (aliph.), 1600 (C = C), 1038 (C = C) cm-1. EI-MS (70 eV), m/z 212 [M]+• (C11H16O4). 1H NMR (CDCl3, 400 MHz): δ 6.62 (1H, m, H-7), 6.53 (1H, m, H-8), 4.56 (1H, broad singlet, H-1), 4.21 (1H, d, J = 5.5 Hz, H-5), 3.96 (1H, dd, J = 1.1; 5.5 Hz, H-4), 3.38 (1H, ddd, J = 0.6; 2.5; 3.5 Hz, H-2), 3.28 (1H, dd, J = 0.6; 2.0; 4.0 Hz, H-3), 1.81 (3H, s, H-10/-11), 1.79 (3H, s, H-10 or H-11). 13C NMR (CDCl3,100 MHz): δ 138.08 (C-9), 132.92 (C-6), 128.34 (C-7), 121.86 (C-8), 73.20 (C-5), 71.00 (C-1 or C-4), 70.90 (C-1 or C-4), 57.81 (C-2), 56.66 (C-3), 26.74 (C-10 or C-11), 18.04 (C-10 or C-11). Panepoxydone (2) - Pale yellow oil. [α]25D -61º(c 0.4, CH2Cl2). IR (KBr) vmax = 3381 (OH), 2974 (aliph.), 2880 (aliph.), 1682 (C = O), 1383 (C-H), 1043 (C-O) cm-1. ESI-MS m/z 209 [M - H]- (C11H14O4). 1H NMR (400 MHz, CDCl3): δ 6.70 (1H, ddd, J = 1.3, 2.5, 3.6 Hz, H-5), 5.29 (1H, qd, J = 1.3, 8.8 Hz, H-7), 5.02 (1H, d, J = 8.8 Hz, H-8), 4.68 (1H, sl, H-4), 3.80 (1H, dq, J = 1.3; 2.5; 3.6 Hz, H-3), 3.46 (1H, dd, J = 1.0; 3.6 Hz, H-2), 1.72 (6H, s, H-10 or H-11). 13C NMR (100 MHz, CDCl3): δ 194.47 (C-1), 138.98 (C-6), 138.26 (C-9), 137.78 (C-5), 123.68 (C-8), 65.35 (C-7), 63.16 (C-4), 57.71 (C-3), 53.88 (C-2), 25.90 (C-10 or C-11), 18.42 (C-10 or C-11). Dihydrohypnophilin (3) - White crystals from CH2Cl2 -Hex. [α]25D +135º (c 0.5, CHCl3). IR (KBr) vmax = 3391 (OH), 2939 (aliph.), 2858 (aliph.), 1666 (C = C), 1458 (aliph.), 1392 (C-H), 1222 (C-O), 1045 (C-O), 957 (C-H), 883 (C-H) cm-1. EI-MS (70 eV), m/z 250 [M]+• (C15H22O3). 1H NMR (400 MHz, CDCl3): δ 5.32 (1H, d, J = 2.2 Hz, H-15a), 5.15 (1H, d, J = 2.2 Hz, H-15b), 4.62 (1H, q, J = 2.2 Hz, H-5), 3.80 (1H, d, J = 8.4 Hz, H-1), 3.47 (1H, d, J = 2.2 Hz, H-6), 2.60 (1H, m, H-9), 2.03 (1H, dd, J = 8.4, 12.0 Hz, H-2), 1.85 (1H, m, H-10α), 1.82 (2H, d, J = 9.0 Hz, H-8), 1.17 (3H, s, H-14), 1.13 (1H, m, J = 12.0 Hz, H-10β), 1.05 (3H, s, H-12), 0.89 (3H, s, H-13). 13C NMR (100 MHz, CDCl3): δ 159.64 (C-4), 112.53 (C-15), 80.96 (C-1), 75.03, (C-7), 74.19, (C-5), 63.73 (C-6), 55.25 (C-2), 47.55 (C-3), 46.36 (C-10), 44.22 (C-11), 34.81 (C-9), 30.74 (C-8), 26.48 (C-12), 19.78 (C-13), 17.51 (C-14). Hypnophilin (4) - Colorless oil. [α]25D -74º (c 0.9, CHCl3). IR (KBr) vmax = 3443 (OH), 2937 (aliph.), 2870 (aliph.), 1728 (C = O), 1636 (C = C), 1464 (C-H), 1375 (C-H), 1115 (C-O) cm-1. EI-MS (70 eV), m/z 248 [M]+• (C15H20O3). 1H NMR (400 MHz, CDCl3): δ 6.13 (1H, s, H-15a), 5.45 (1H, s, H-15b), 3.87 (1H, d, J = 9.0 Hz, H-1), 3.43 (1H, s, H-6), 2.64 (1H, m, H-9), 2.14 (1H, dd, J = 9.0, 12.0 Hz, H-2), 1.95 (2H, m, H-8), 1.90 (1H, m, H-10α), 1.30 (3H, s, H-14), 1.23 (1H, m, H-10β), 1.07 (3H, s, H-12), 0.89 (3H, s, H-13). 13C NMR (100 MHz, CDCl3): δ 197.60 (C-5), 153.73 (C-4), 121.61 (C-15), 81.23 (C-1), 76.04 (C-7), 61.14 (C-6), 56.13 (C-2), 46.12 (C-10), 45.54 (C-3), 44.14 (C-11), 34.52 (C-9); 30.75 (C-8), 26.43 (C-12), 19.66 (C-13), 17.75 (C-14). Cell preparations - Venous blood from healthy adult volunteers was collected in heparinized tubes and centrifuged over a Ficoll-Hypaque cushion (Histopaque, Sigma, St Louis, MO). Peripheral blood mononuclear cells (PBMC) were collected from the Ficoll/Hypaque interphase and washed three times in RPMI prior to further processing. The cell suspensions were adjusted to 1.5 x 106 cells/ml. Viability assays - PBMC viability was determined using a modification of the protocol described by Souza-Fagundes et al. (2002). Briefly, the cells were treated with pure compounds at different concentrations for 72 h at 37ºC in a 95% relative humidity atmosphere with 5% CO2. After this period, the cells were incubated for 10 min with trypan blue (0.4% in NaCl 0.9%). In the flow cytometry experiments, trypan blue fluorescence was measured in the FL2 channel. Fluorescence intensity was used to discriminate viable (non-fluorescent) and non-viable (fluorescent) lymphocytes. Controls with 0.1% DMSO run in parallel did not demonstrate any differences in cell viability (data not shown). PBMC culture - All cultures were performed in RPMI-1640 medium (GIBCO, Grand Island, NY), supplemented with 5% (v/v) heat-inactivated, pooled human sera type AB (Flow Laboratories, Royaune, UN) and L-glutamine (2 mM, GIBCO, Grand Island, NY). An antibiotic/antimicotic solution containing 1,000 U/ml penicillin, 1,000 µg/ml streptomycin and 25 µg/ml fungisone (SIGMA, St. Louis, MO) was added to control fungal and bacterial contamination. Lymphocyte proliferation assay - In vitro cellular proliferation (blastogenesis) was assessed as previously described (Gazzinelli et al. 1983). Briefly, 1 x 106 cells/ml (1.5 x 105 cells per well) were cultured in complete RPMI-1640 in flat-bottomed microtiter plates (Costar, tissue culture treated polystyrene # 3512, Corning, NY, USA). The cultures were stimulated with 2.5 µg/ml of PHA (SIGMA, St Louis, MO) and incubated for 72 h at 37°C in a humidified atmosphere containing 5% CO2. The cell proliferation and viability were determined using an MTT (methyl thiazolyl tetrazolium)-based colorimetric assay (Jiang & Xu 2003). The results were expressed as percent inhibition of the lymphocyte proliferation in relation to the control (without drugs). Assay with recombinant TR from T. cruzi - The recombinant TR assay was performed according to Hamilton et al. (2003) in 96-well plates (Costar 9017, Corning, USA) using Hepes buffer (40 mM pH 7.5) with 1 mM EDTA. Each assay well (250 µl) contained enzyme (6 mU), trypanothione (0.25 nmol) and NADPH (50 nmol). The extracts, fractions or pure compounds were added to the above mixture and incubated at 30ºC during 30 min. After this period, 17.5 nmol of DTNB [5,5'-dithiobis(2-nitrobenzoic acid), Ellman's reagent] was added, and the absorbance (Abs) was measured at 412 nm in the kinetic mode every 10 s for 5 min. The resulting slope of the δAbs/δt plot is proportional to DTNB reduction and the enzyme activity (Hamilton et al. 2003). The enzyme inhibition was calculated as the ratio between (δAbs/δt) of the experimental wells and that of the controls without drug, that is, percent inhibition = (1 - (δAbs)exp/(δAbs)contr ) x 100. Assay with T. cruzi amastigotes - This assay was performed as described previously by Buckner et al. (1996) using a T. cruzi (Tulahuen strain) expressing the Escherichia coli β-galactosidase gene. Infective trypomastigote forms were obtained through culture in monolayers of mouse L929 fibroblasts in RPMI-1640 medium (pH 7.2-7.4) without phenol red (Gibco BRL) plus 10% fetal bovine serum, 2mM glutamine and 40 µg/ml gentamycin. For the bioassay, 4,000 L929 cells in 80 µl of supplemented medium were added to each well of a 96-well microtiter plate. After an overnight incubation, 40,000 trypomastigotes in 20 µl were added to the cells and incubated for 2 h. The medium containing extracelullar parasites was replaced with 200 µl of fresh medium and the plate was incubated for an additional 48 h to establish the infection. The medium was then replaced with solutions of the compounds at different concentrations in fresh medium (200 µl), and the plate was incubated for 96 h. After this period, 50 µl of 500 µM chlorophenol red glycoside in 0.5% Nonidet P40 was added to each well, and the plate was incubated for 24 h, after which the absorbance at 570 nm was measured. Controls with uninfected cells, untreated infected cells, and medium only were run in parallel. The results are expressed as percent reduction of the absorbance in the experimental wells in comparison with the control wells with untreated infected cells (percent inhibition = (1 - Absexp/Abscontr) x 100). Triplicates were run in the same plate, and the experiments were repeated at least once. RESULTS L. strigosus (Schwein) Fr. was grown in MEC medium, and the culture extracted with ethyl acetate yielded 2.29 g of a crude extract that was able to inhibit TR activity by 100% at 20 µg/ml. In order to identify the active components of the extract, 100 µg of the crude extract was fractionated in an analytical HPLC column, and the effluent was collected in a 96-well microplate (250 µl/well) while recording the UV chromatogram (at 210 nm). After solvent elimination, the dry films in the plate were dissolved with the assay buffer, and the solutions were subjected to the TR inhibition assay. The results of this experiment are presented as an overlay plot of the chromatogram and as the percentage of TR inhibition (Fig. 1), clearly showing that the inhibitory activity was concentrated on compounds eluting around 13 and 19 min. The HPLC method was then scaled up to isolate the compounds in sufficient amounts for structural elucidation and IC50 determinations. Four compounds were isolated, either by crystallization of the collected fractions or after further chromatographic steps, as described in the Materials and Methods section. Based on the analysis of their UV, IV, Mass, and NMR spectra and on comparison with data reported in the literature, the compounds were identified as neopanepoxydol (1), panepoxydone (2), dihydrohypnophilin (3) and hypnophilin (4), as shown in Fig. 2. Detailed analysis of the HMQC and HMBC NMR data allowed the unambiguous assignments of C-12 and C-14 chemical shifts in dihydrohypnophilin and hypnophilin, requiring the correction of those proposed by Abate and Abraham (1994). Thus, the attributions for C-12/C-14 in dihydrohypnophilin should be corrected from 17.5/26.5 δ to 26.48/17.51 δ, and in hypnophilin from 17.6/26.4 δ to 26.43/17.45 δ. All pure compounds were tested at 20 µg/ml in the TR, amastigote, and PBMC assays (Table I). The results show that panepoxydone and hypnophilin were active in all bioassays while neopanepoxydol and dihydrohypnophilin were inactive. Dose response experiments with the active compounds confirmed these preliminary results, with hypnophilin and panepoxydone disclosing IC50 values around 1 and 39 µM, respectively (Table II). Hypnophilin was also more active than panepoxydone against intracellular amastigotes, having an IC50 value of 2.5 µM. Panepoxydone, however, was more effective in inhibiting PBMC proliferation, having an IC50 of 1.3 µM. From the experimental data used to construct the dose response curves (data not shown), it was estimated that 4 µM hypnophilin is able to block 100% of TR activity and inhibit 80% of the growth of intracellular amastigotes, with only minimal inhibitory activity (20%) on the proliferation of PHA-stimulated PBMC (Fig. 3). According to the data, at this concentration, panepoxydone would be almost inactive against amastigotes and still present some toxicity to PBMC. To investigate this hypothesis, the impact of panepoxydone and hypnophilin at 4 µM on lymphocyte and monocyte subpopulations of the PBMC was evaluated. The results (Fig. 4a,b) show that panepoxydone induced cell death on the lymphocyte and monocyte populations compared with PBMC control cultures while, as expected, hypnophilin was not cytotoxic. These results were corroborated by quantitative analyses by flow cytometry (Fig. 4c), which showed that panepoxydone reduced the lymphocytes population by 20%, while hypnophilin showed only a 5% reduction, which is similar to the control culture without drug. Similar results were observed for the monocytes population, where panepoxydone significantly induced cell death (21%), while the percentage of death caused by hypnophilin mirrored the control without treatment (11%). DISCUSSION A bioassay-guided procedure based on the inhibition of TR was used to detect and isolate neopanepoxydol (1), panepoxydone (2), dihydrohypnophilin (3) and hypnophilin (4) from L. strigosus. Our results confirm the efficiency of the method based on analytical HPLC and off-line bioassay to identify the retention time of the active components (panepoxydone and hypnophilin) present in the extract, facilitating their isolation. Hypnophilin (4) was previously isolated from the culture of Pleurotellus hypnophilus (Gianetti et al. 1986), Lentinus critinus (Abate & Abraham 1994) and L. connatus (Rukachaisirikul et al. 2005). Dihydrohypnophilin (3) was isolated from a culture of L. crinitus (Abate & Abraham 1994) and L. conatus (Rukachaisirikul et al. 2005). Panepoxydone and neopanepoxydol were obtained from fermentations of Panus rudis and Panus conchatus (Kis et al. 1970), while panepoxydone was isolated with hypnophilin from cultures of the L. connatus (Rukachaisirikul et al. 2005) and L. crinitus (Erkel et al. 1996). Panepoxydone and hypnophilin inhibited TR in the same concentration range of a series of synthesized polyaminoguanidines and polyaminobiguanides (IC50 values from 0.95 to 69.47 µM) (Bi et al. 2006). Under our assay conditions, hypnophilin inhibits TR in the low micromolar range (IC50 = 1 µM), making it about three times more active than the reference compound clomipramine (IC50 = 3 µM), a known inhibitor of TR. It is also more active than the macrocyclic spermidine alkaloid lunarine (IC50 = 65 µM, Hamilton et al. 2005), which was isolated from Lunaria biennis. These results indicate that hypnophilin can be considered a good starting point for a medicinal chemistry program aimed at determining derivatives with higher TR inhibitory activity. Concerning the mechanism by which panepoxydone (2) and hypnophilin (4) inhibit TR, we speculate that it may be similar to that proposed for the alkaloid lunarine (Hamilton et al. 2005), in which the presence of conjugated carbonyl groups make it susceptible to nucleophilic attack by the thiol groups present in the active site of TR. The lack of activity by neopanepoxydol (1) and dihydrohypnophilin (3), which are closely related to two and four but do not carry the conjugated carbonyl moiety, reinforces this hypothesis. Besides its activity in the biochemical assay, hypnophilin was also active against the intracellular amastigotes of the Tulahuen strain of T. cruzi infecting L929 cells. Inhibition of the multiplication of T. cruzi intracellular amastigotes is clinically relevant because these are the proliferating forms of the parasite. These parasites mature into trypomastigote forms that cause the rupture of the host cells, thus promoting tissue damage in the infected organs, especially the esophagus, colon and heart of chronic patients of Chagas disease. Although circulating parasites cannot be observed by blood inspection in this phase, T. cruzi antigens may trigger the immune system by multiple mechanisms resulting in inflammation and fibrosis. An inflammatory response may result in accumulation and activation of monocytes in addition to increasing the concentration of macrophage-derived cytokines in the plasma (Aliberti et al. 1996). These cytokines can promote an immediate response and also affect organs and tissues. There is a general consensus that the host immune response plays a pivotal role in the adverse events caused upon the massive antigen release triggered by trypanocidal agents during treatment (Sathler-Avelar et al. 2006). With this in mind, our results show that hypnophilin display a non-cytotoxic antiproliferative activity on PBMC stimulated with PHA, suggesting that, besides its trypanocidal activity, hypnophilin can eventually reduce tissue damage by interfering with the proliferation of cells involved with the immune response. It is important to mention that the currently available treatment for Chagas disease with benznidazole or nifurtimox is effective mainly during the acute phase of infection, with little effectiveness during chronic disease (Tanowitz et al. 1992, Prata 2001). Furthermore, benznidazole and nifurtimox are more effective against the blood forms observed in the acute phase than the intracellular amastigote forms of the chronic phase (Muelas-Serrano et al. 2002). It must be emphasized that, with control of natural transmission in many areas afflicted by Chagas disease, new cases are episodic and the majority of the 18 million patients with T. cruzi are currently in the chronic phase (Prata 2001). In this connection, our findings indicate that hypnophilin is a lead candidate for development of new drugs to treat chronic Chagas disease. Additional studies are needed to characterize the molecular basis of hypnophilin-induced immunomodulatory activity. We speculate that it may involve the up-regulation of IL-10 synthesis, as is the case with benznidazole (Pascutti et al. 2004), or the engagement of other cytokines and/or apoptosis mechanisms that would allow the effective parasite killing in the absence of immunomediated tissue damage. Two metabolites, panepoxydone and hypnophilin, with significant TR inhibitory activity were isolated from L. strigosus, a tropical basidiomycete investigated for the first time. Hypnophilin disclosed significant anti-T. cruzi activity associated with minor non-cytotoxic immunomodulatory activity on human leukocytes, suggesting that it is a good lead candidate for developing new drugs against Chagas disease. ACKNOWLEDGEMENTS To Rosana Alves, Rodrigo Leite and Aline Vaz for their technical assistance. To Prof. Alan Fairlamb (Univ. of Dundee, Scotland) for for providing the recombinant trypanothione reductase, and to Prof. Frederick Buckner (University of Washington, USA) for the Tulahuen strain of T. cruzi expressing the E. coli β-galactosidase gene. REFERENCES
Copyright 2008 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc08045f3.jpg] [oc08045t1.jpg] [oc08045t2.jpg] [oc08045f4.jpg] [oc08045f2.jpg] [oc08045f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}