 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
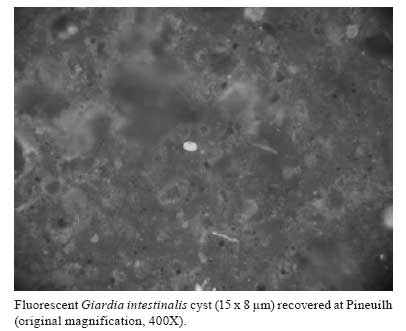
Memórias do Instituto Oswaldo Cruz, Vol. 103, No. 3, May 2008, pp. 298-300 New finding of Giardia intestinalis (Eukaryote, Metamonad) in Old World archaeological site using immunofluorescence and enzyme-linked immunosorbent assays Matthieu Le BaillyI, II, +; Marcelo LC GonçalvesIII, IV; Stéphanie Harter-LailheugueI; Frédéric ProdéoV; Adauto AraujoIV; Françoise BouchetI IUniversité de Reims, EA 3795, UFR de Pharmacie, Laboratoire de Paléoparasitologie, CNRS UMR 5197, 51, rue Cognacq-Jay, 51096 Reims, France Financial support: CNRS, INRAP, Ile-de-France region, Capes/Cofecub, CNPq, Millipore®, Fumouze, Meridian diagnostic Received 27 February 2008 Code Number: oc08051 ABSTRACT In this study, nine organic sediment samples from a medieval archaeological site at Pineuilh, France, were examined for Giardia intestinalis using two commercially available immunological kits [enzyme-linked immuno sorbent and immunofluorescence (IFA) assays]. Both techniques detected G. intestinalis in one sample, dated to 1,000 Anno Domini. This is the first time IFA was successfully used to detect protozoa in Old World archaeological samples. Such immunological techniques offer important perspectives concerning ancient protozoa detection and identification. Key words: paleoparasitology - ELISA - IFA - Giardia intestinalis Detection of protozoa in ancient faeces (coprolites) still poses a challenge to scientists interested in parasitic disease and host-parasite interaction evolution, because protozoan cysts are less resistant to taphonomic process than helminth eggs (Gonçalves et al. 2005). However, by applying immunological techniques, it is possible to overcome this limit. Many results published since the 1980s prove the reliability of immunology to detect protozoan infections in ancient populations. These findings concern blood protozoa such as Trypanosoma cruzi (Fornaciari et al. 1992) or Plasmodium falciparum (Miller et al. 1994, Cerutti et al. 1999), and intestinal protozoa such as Entamoeba histolytica (Fouant et al. 1982, Gonçalves et al. 2004, Le Bailly & Bouchet 2006, Le Bailly et al. 2006). Giardia intestinalis has also been found in archaeological samples mainly coming from New World settings. Faulkner et al. (1989) detected G. intestinalis in a coprolite from the Big Bone Cave site, Tennessee, USA, dated to the III century BC, using immunofluorescence assay (IFA). Allison et al. (1999) then Ortega and Bonavia (2003) found both G. intestinalis and Cryptosporidium parvum, using IFA and enzyme-linked immunosorbent assay (ELISA) on coprolite samples from pre-Columbian (XI century BC to XV century AD) and Peruvian mummies (XXIV century BC to IX century AD). Gonçalves et al. (2002) detected G. intestinalis in archaeological samples, coprolites and sediments, from different sites and different periods of time, using ELISA technique. So far, the oldest finding of G. intestinalis in archaeological sites was reported by Le Bailly (2005), examining samples from the European medieval site of Chevennez (VII-IX centuries AD), Switzerland, by ELISA. Although intestinal protozoan infections have been recorded in different New World archaeological sites, little is know about their ancient distribution in Old World. MATERIALS AND METHODS To compensate the lack of study on giardiasis in Old World and to continue developing the use of immunology in paleoparasitology, nine sediment samples (S1 to S9) coming from humid habitation layers from "La Mothe", a French medieval site (X-XI centuries AD), were studied for the first time to test the presence of G. intestinalis using two commercially available immunological assays. The first, the MeriFluor Crypto & Giardia test (Meridian Bioscience, Cincinnati, OH, USA), is an IFA to detect Cryptosporidium sp. and G. intestinalis in actual stool specimens, based on monoclonal antibodies (Garcia et al. 1987, Aldeen et al. 1995). The second, Giardia II test (TechLab, Inc., Blacksburg, VA, USA), is an immune-enzymatic kit which allows the qualitative detection of a cyst wall antigen of G. intestinalis in faecal specimens (Aldeen 1998, Boone et al. 1999). The regular process we apply for archaeological sample rehydration uses a trisodic phosphate and glycerinated water solution in which some drops of formalin are added to avoid fungi and bacteria development (Le Bailly 2005). To avoid modifications of antibody/antigen link reactions due to the formalin, archaeological sample were rehydrated only using ultrapure water (Millipore®) and stored in a refrigerator at 2-5ºC to prevent micro-organism development. Aliquots from the rehydrated samples were used in the immunological assays. Both direct IFA and ELISA were performed according to the manufacturer's instructions. RESULTS AND DISCUSSION Only one sample (S1) was positive for G. intestinalis antigen by ELISA and direct IFA. Under UV microscopy, S1 yielded fluorescent ovoid elements with size of 15 x 8 µm. These elements were identified as G. intestinalis cysts (Figure). Other samples were negative for G. intestinalis in both tests. The nine samples were negative for Cryptosporidium sp. antigen detection by IFA. The presence of G. intestinalis in S1 can be attributed to human or animal faecal pollution. However, the presence of the human specific nematode Enterobius sp. eggs in the sample, identified during a previous regular microscopically analysis, proves the presence of a faecal pollution of human origin. G. intestinalis is a faecal-oral transmitted flagellate protozoan transmitted by cyst ingestion after faecal food contamination (Mehlhorn 2001). Trophozoites are liberated in the small intestine after excystation, and attach to the small intestine epithelium, causing digestive symptoms of variable importance, among which diarrhoea is the main associated (Nozais et al. 1996, Gentilini 1999, Acha & Szyfres 2005). Although the specificity of the ELISA Giardia II test had been evaluated using faecal specimens containing many other parasites, including E. histolytica, Ascaris lumbricoides, Trichuris trichiura, and ancylostomidae, false-positive results can not totally be excluded. IFA, on the other hand, has the advantage to allow direct observation of cysts under optical microscopy. According to the manufacturers, the specificity of the IFA is estimated to be over 99% (Johnston et al. 2003), with no cross-reaction with E. histolytica, Escherichia coli, Entamoeba hartmanii, Endolimax nana, and Chilomastix mesnili. This study presents the first detection of ancient protozoan cysts using IFA in the Old World. The remarkable conservation conditions of the site and the presence of a humid and anaerobic environment favoured the preservation of parasitological remains and the discovery of intact cysts. The presence of G. intestinalis in S1 could reflect some sanitation problems and bad human and animal excrement management. This work also completes the parasitological records for the X-XI centuries AD medieval period, so completing a chronological hiatus in the European record of giardiasis. G. intestinalis cysts were recorded only in small numbers and in relatively recent archaeological sites in the Old World (VII-IX centuries AD) (Le Bailly 2005). This may contribute to debate the origin of the parasite strain in the New World, where G. intestinalis infection was recorded in Peruvian mummies dated to XXIV century BC (Ortega & Bonavia 2003). The use of IFA is particularly important in paleoparasitology, allowing the confirmation of some ELISA results, and thus decreasing false positive risks. Moreover, because of their specific ability in identifying parasitic remains, immunological tools present a major field in ancient parasite diagnostic. Systematic utilization of immunology should increase the species diversity detected in archaeological contexts, and will allow mapping the parasite historical records, clarifying ancient human migration pathways and human-parasite co-evolution. REFERENCES
Copyright 2008 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc08051f1.jpg] |
| |||||||||

{kind=link}