 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 103, No. 4, June, 2008, pp. 313-325 REVIEW Electron microscopy of trypanosomes - A historical view Wanderley de SouzaI, II ILaboratório
de Ultraestrutura Celular Hertha Meyer, Instituto de Biofísica Carlos
Chagas Filho, Universidade Federal do Rio de Janeiro, CCS-Bloco G, Ilha do Fundão,
21941-900 Rio de Janeiro, RJ, Brasil Financial support: During the course of this study, work performed in the author's laboratory has been supported by grants from CNPq, Finep and Faperj Received: 12 June
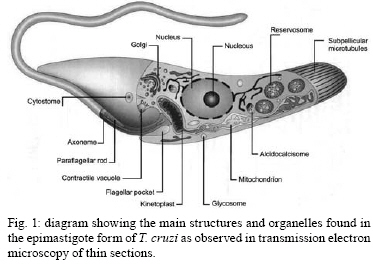
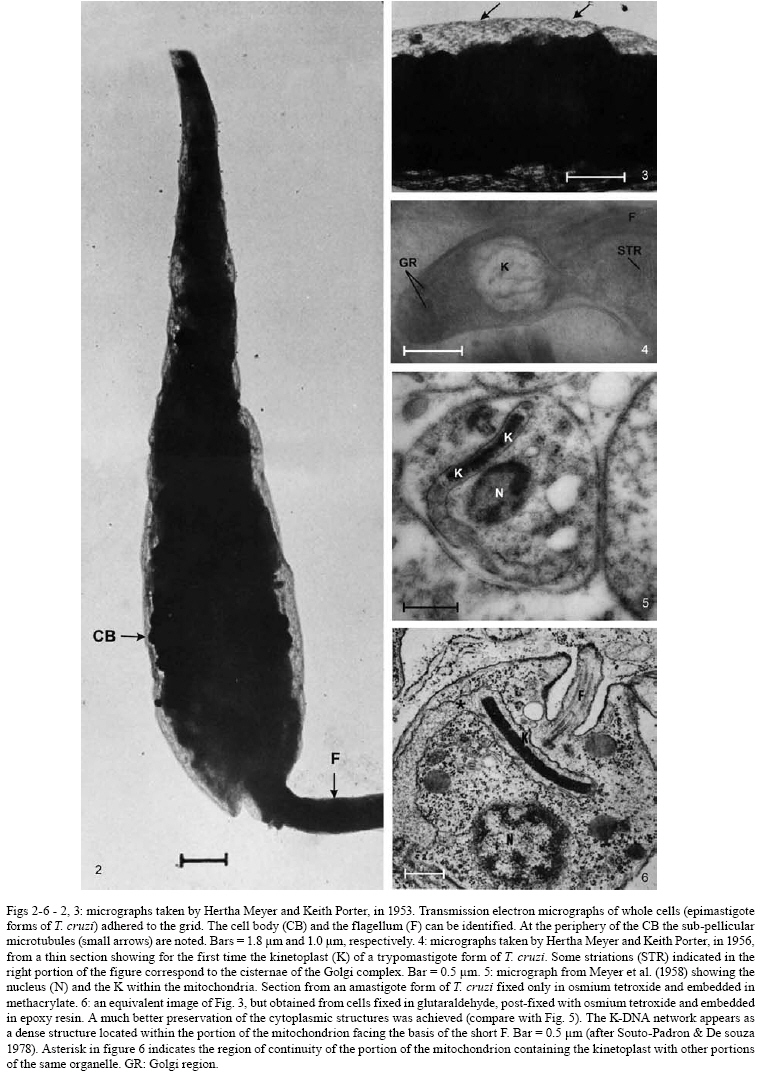
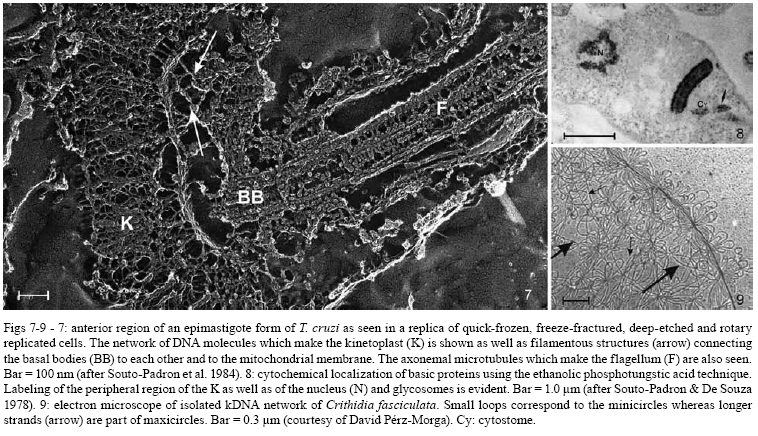
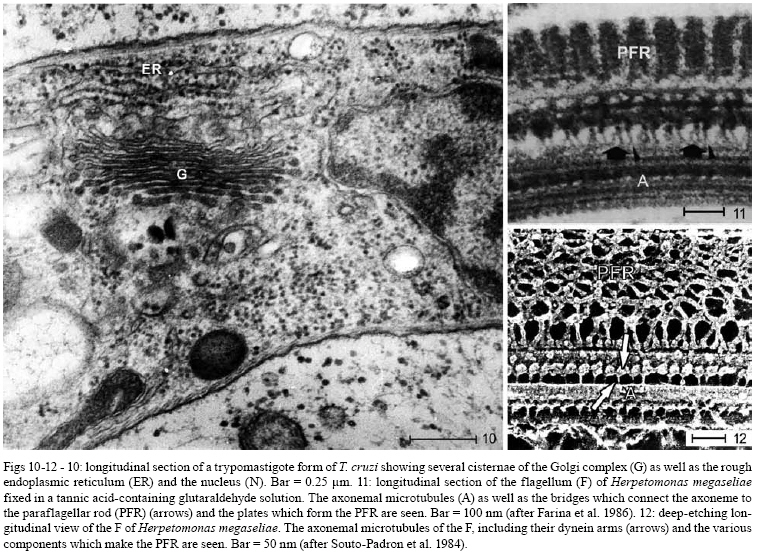
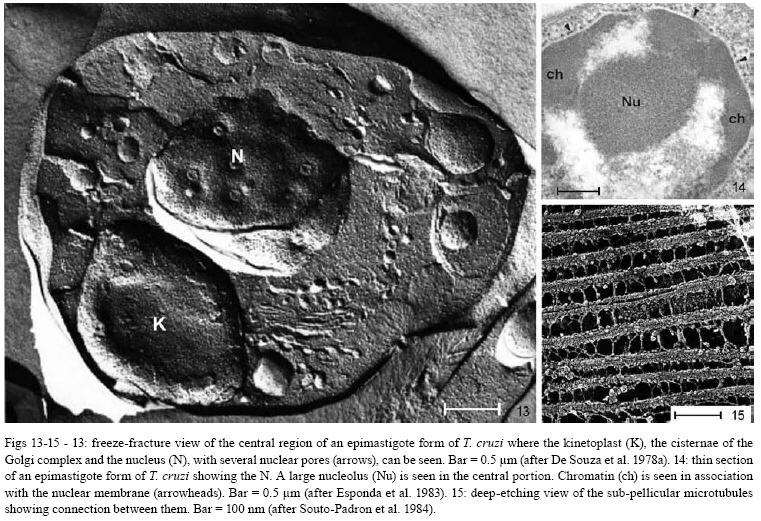
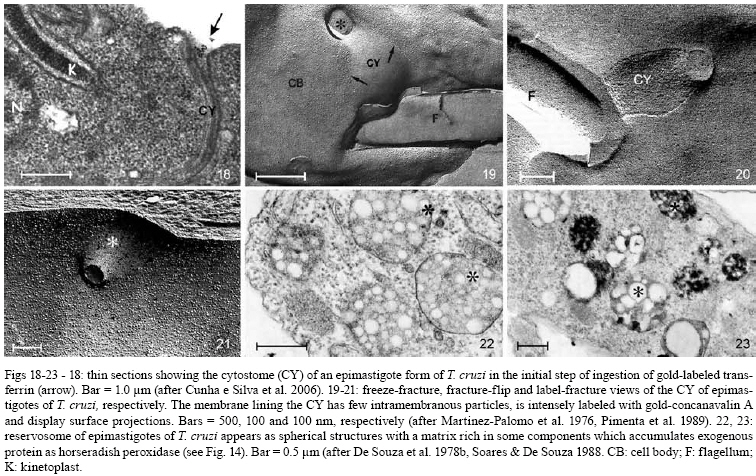
2008 Code Number: oc08055 ABSTRACT Since the discovery of the electron microscope and the development of the initial techniques for the processing of biological samples for electron microscopy, the protozoan Trypanosoma cruzi has been the subject of intense investigation. This review analyzes the results obtained by observation of whole trypanosomes as well as thin sections and replicas using several microscopic approaches. Micrographs detailing the appearance of T. cruzi using several methods illustrate the evolution of electron microscopic techniques as well as its contribution to understanding the structural organization of the protozoan. Key words: electron microscopy - ultrastructure - trypanosomes - Trypanosoma cruzi Introduction and aims There are few cell types that have been so intensely analyzed using all microscopy techniques available as trypanosomes. The visualization of living trypanosomes by light microscopy (LM) impressed the first observers because of the motility of the parasites, as they moved both forward and backward, and their constant contracting and wriggling movements seemed to be due to an undulating membrane of the body and to the flagellum. Probably due to the fact that some trypanosomes are the causative agents of highly relevant diseases, such as African sleeping sickness (Trypanosoma brucei), American Chagas disease (Trypanosoma cruzi), and several clinical forms of leishmaniasis seen throughout the world (caused by different Leishmania species), this group of eukaryotic organisms has been the subject of intense investigation. According to the World Health Organization, these protozoa infect about 37 million people. Since almost all available electron microscopy (EM) techniques have been used to analyze the structural organization of these organisms, trypanosomes constitute an excellent model to demonstrate the evolution of the biological EM over the past 60 years. In this paper, we will review the main contributions made by microscopic techniques to our present day knowledge on the structural organization of trypanosmatids, giving more emphasis on T. cruzi. Members of the Trypanosomatidae family undergo a complex life cycle. In the case of T. cruzi this cycle involves both invertebrate and vertebrate hosts (review in De Souza 2002). The cycle in the invertebrate host starts when the triatomine ingests blood from infected vertebrate hosts (either humans or wild mammals such as marsupials and armadillos) containing trypomastigote forms. In the stomach of the triatomine, the trypomastigote forms transform into rounded spheromastigotes and subsequently into epimastigotes. These proliferate in the midgut, attach themselves to the intestinal cells and then transform into the highly infective metacyclic trypomastigotes that are released immediately after blood meals in the feces as well as in the insect urine. These forms, when inoculated into a vertebrate host, begin a new cycle. In mammals, the cycle starts with the penetration of metacyclic trypomastigotes into cells found in the skin through a process of endocytosis. Following invasion, the trypomastigote forms found within an endocytic vacuole, known as the parasitophorous vacuole, gradually transform into the rounded amastigote forms and almost simultaneously disrupt the membrane lining the vacuole. Then, the amastigote forms start to divide in direct contact with cytoplasmic structures. After several division cycles, a large number of amastigotes can be found within the host cell. Then, due to not yet characterized stimuli, the amastigotes transform into trypomastigotes, rupturing the host cell membrane, and many trypomastigotes are released into the intercellular space. These forms are then able to infect new cells and amplify the infection in the vertebrate host. This cycle can be reproduced in vitro in cell cultures. Most of the studies on T. cruzi have been performed with the epimastigote form, the proliferative developmental stage found in the intestine of the invertebrate host, which can be easily grown in axenic cultures. For a better understanding of the description of the various structures and organelles found in T. cruzi, we present a schematic view of the fine structure of the epimastigote form based on our current knowledge (Fig. 1). Electron microscopy in Biology in the 1940s and 1950s Although the transmission electron microscope was invented by Ernest Ruska in 1934, the first interesting results with biological samples were not obtained until the 1940s. In a classical work, Porter et al. (1945) reported the results of microscopically observing whole cells maintained in tissue cultures (TC), a condition where some cells spread well, forming thin cytoplasmic projections. Such images revealed details of the cell organization and the clear identification of the mitochondria and the description of a membranous network later on known as the endoplasmic reticulum. At this time, images were also obtained by the examination of platinum-palladium replicas of the cells in the electron microscope. Structures visualized in whole cells The trypanosome was also a 'personage' in the early period of EM history. Around 1948, Carlos Chagas Filho visited the Rockefeller Institute in New York and saw the images obtained by Porter and colleagues in mammalian cells maintained in vitro. When he returned to Brazil, he encouraged Hertha Meyer to observe trypanosomes using the same approach. In 1950, Meyer began studying T. cruzi with the electron microscope in the laboratory of Keith Porter. At that time, it was not yet possible to obtain thin sections, although many groups were trying to adapt a conventional microtome for cutting thinner sections. The first attempts used TC infected with T. cruzi or even with culture forms of the protozoan. The cells were fixed with osmium tetroxide, washed several times and then one drop of the cell suspension was placed on the copper grids previously coated with Parlodion and allowed to dry in dust-protected dishes. The first results obtained were rather disappointing. In spite of the small size of the flagellates, which are much smaller than a mammalian cell, nothing could be seen of their inner structure (Figs2, 3). A good electron penetration was only obtained at the periphery, and the cytoplasm showed a fine striation in parallel array, corresponding to the now well-known sub-pellicular microtubules (Meyer & Porter 1954). Very dense spherical bodies could be seen in the body portion of the parasite. The whole body seemed to be covered by a thin sheath. Trypsinization or prolonged fixation with osmium tetroxide destroyed the fine striation in the periphery of the cell and also the fiber bundle that composed the flagellum, suggesting that the cell body and the flagellum had their own sheath that simply adhered to one another. The first thin sections It was not until the 1950s that the first thin sections were obtained. First, it was necessary to develop ultramicrotomes with the required stability and ability to cut thin (nm range) sections. Initial attempts were made in the 1940s by several groups, including an important work published by Pease and Baker (1948). An important achievement was the development of glass knives by Latta and Hartman (1950). However, it was the seminal work of Porter and Blum (1953) that developed equipment that worked well and became the prototype of a whole series of ultramicrotomes known as the Porter and Blum microtomes. One of the first ultramicrotomes made at the Rockefeller Institute was given to Hertha Meyer from Keith Porter, and was intensely used to obtain thin sections of pathogenic protozoa. It is important to mention that a fundamental step for the obtaining of quality thin sections was the development of the embedding media, which could resist an electron beam. Methacrylate was introduced in 1949 (Newman et al. 1949), araldite in 1958 (Glauert & Glauert 1958), and epoxy resins in 1961 (Luft 1961). The latter mediums gave excellent results allowing thin sections to be easily obtained from the blocks formed and where the cell structures were well stained (or contrasted) with metals such as uranyl acetated (Watson 1958) and lead citrate (Reynolds 1963), greatly improving the quality of the resulting images. From these advances in techniques, the internal structures of T. cruzi were unveiled. Below is a list of structures that were identified in T. cruzi during the first phase of biological EM of thin sections when the samples were primarily fixed in osmium tetroxide. The kinetoplast By 1956, Hertha Meyer obtained the first thin sections of T. cruzi in Keith Porter's laboratory, leading to a publication in 1958 (Meyer et al. 1958). The images could clearly identify a dark, electron dense and slightly bent inclusion that would correspond to a structure known at that time, based on the observation by LM of stained cells, as the kinetonucleus (Fig. 4). This structure is now known as the kinetoplast. It is situated proximal to the nucleus and its shape and structural organization varies according to the protozoan developmental stage. With higher magnification, it could be seen that the kinetoplast was represented by a vacuole-like space that contained an electron dense material. In longitudinal sections, this inclusion showed a lamellar, almost spiral-like structure that was not separated by a membrane from the surrounding electron transparent space of the vacuole. At the same time, Steinert et al. (1958) demonstrated that the kinetoplast incorporates [3H] thymidine into the fibrillar structure, thus confirming the presence of DNA. Further studies using electron microscope autoradiography, performed by Burton and Dusanic (1968) as well as Anderson and Hill (1969), showed the dynamics of 3H thymidine incorporation during kinetoplast replication. The remarkable molecular configuration of the DNA contained in the kinetoplast was only revealed in the late 1960s and early 1970s (Riou & Yot 1967, review in Shapiro & Englund 1995, De Souza & Cavalcanti 2008). The vacuole containing the fibrillar structure, now known as the kinetoplast-DNA network, continues directly into the large canal system made of septae or cristae, similar to those described by George Palade and others as typical for mitochondria in tissue cells. What Meyer and co-workers described at that time has been confirmed. It is currently known that Trypanosomatids possess a unique and highly ramified mitochondria (Paulin 1975). Subsequently, it was shown that the dense structure is made of a special type of DNA known as the kinetoplast DNA (K-DNA). The first images obtained using thin sections (Fig. 5) clearly revealed the organization of the K-DNA (Meyer et al. 1958), and showed that it was located in a specialized portion of the mitochondrion (Fig. 5), as clearly indicated in Fig. 6. The K-DNA is located within the mitochondrial matrix, perpendicular to the axis of the flagellum. In some trypanosomatids, the position of the kinetoplast relative to the nucleus changes during the life cycle. However, the kinetoplast is always located proximal to the basal body. Despite the close proximity of these structures, evidences that the kinetoplast is physically linked to the basal body did not arise until the 1980's (Fig. 7) (Souto-Padron et al. 1984). Recently, the presence of a set of filaments connecting the kinetoplast DNA to the basal body was demonstrated, providing an explanation for the spatial position and segregation of the trypanosome mitochondrial genome (Ogbadoiyi et al. 2003). Due to this connection, the position of the kinetoplast defines the cellular region in which the basal body is located and consequently the origin of the flagellum. Recently, a protein designated as p166 was identified and shown to be located between the K-DNA disk and the flagellar body (Zhao et al. 2008). In addition to DNA, EM cytochemistry, using the ethanolic phosphotungstic acid technique and ammoniacal silver (Souto-Padron & De Souza 1978), showed the presence of basic proteins in the kinetoplast (Fig. 8) and suggested that these proteins could neutralize the negatively charged DNA molecules that are in close contact within the kinetoplast. Currently, biochemical and molecular evidences have confirmed the presence of basic proteins in the kinetoplast, indicating that histone-H1 like proteins participate in the K-DNA condensation in Crithidia fasciculata and T. cruzi (Xu & Ray 1993, Caval- canti et al. 2004). The K-DNA represents about 30% of the total cellular DNA and differs from nuclear DNA in buoyant density, base ratio and degree of renaturation. Moreover, unlike any other DNA in nature, the K-DNA of trypanosomatids is composed of circular molecules, which are topologically relaxed and interlocked to form a single network. Two types of DNA rings are present in the kinetoplast, the minicircles and the maxicircles (Fig. 9). There are several thousand minicircles, which range in size from about 0.5 to 2.5 kb (depending on the species) and a few dozen maxicircles, usually varying between 20-40 kb (review in Shapiro & Englund 1995). These structures encode guide RNAs, which modify the maxi-circle transcripts by extensive uridylate insertion or deletion in a process known as RNA editing. The maxicircles are structurally and functionally analogous to the mitochondrial DNA from higher eukaryotes, encoding rRNAs and subunits of the respiratory complexes. Fluorescence microscopy played a fundamental role in the understanding of the replication of K-DNA. Using bromodeoxyuridine to label replicating free minicircles and the TdT (terminal deoxynucleotidyl transferase) technique to reveal the already replicated gaped circles, Liu & Englund (2007) showed that replication occurs in approximate synchrony with the nuclear S phase and requires a repertoire of molecules, including type II topoisomerases, DNA polymerases, universal minicircle sequence binding proteins, primases and ribonucleases. Recent reviews offer further data on the kinetoplast (Shapiro & Englund 1995, Lukes et al. 2002, Liu et al. 2005). The Golgi complex (STR structure) The first thin sections showed the presence of some parallel striations localized between the kinetoplast and the nucleus and proximal to the region of emergence of the flagellum (Fig. 4). With subsequent improvements in techniques, it became clear that the striations correspond to what was being described in mammalian cells as parallel cisternae or sacculae, which form the structure known as the Golgi complex. The Golgi structural organization in trypanosomes is similar to that described in other cells (Fig. 10). The Golgi complex was isolated and further characterized using biochemical methods. Using gold-labeled lectins to reveal sugar-containing molecules, the organelle was labeled and found to be involved in protein glycosylation as reported for other eukaryotic cells. Rab 7, a small GTPase involved in membrane trafficking, was also detected in the Golgi complex of trypanosomes (Araripe et al. 2004). Basal body-flagellum complex A typical basal body located at the basis of the unique flagellum was observed in the first thin sections examined of all T. cruzi developmental stages. Even in the so-called amastigote form a short flagellum was observed. The first EM studies revealed the presence of an intriguing structure in the flagellum of trypanosomatids that was designated as the paraxial or paraflagellar rod (PFR) due to its localization. This structure is much less developed in those species that contain a bacteria endosymbiont (Gadelha et al. 2005). Here, it is clear how new EM techniques are powerful enough to reveal new sub-structures. For instance, the use of tannic acid in association with glutaraldehyde significantly improves the preservation of sub-structures (Fig. 11). Subsequently, the use of quick-freezing, freeze-fracture, deep-etching and rotary replication (Souto-Padron et al. 1984, Farina et al. 1986) allowed the observation of several structures that were not visualized in thin sections (Fig. 12). It was seen that the PFR is made of a complex array of filaments linked to the axoneme. Two regions, designated proximal and distal, were identified in the PFR, and are formed by two and by several plates, respectively. The plates are formed by an association of 25 nm and 7 nm thick filaments that are oriented at 50° angle in relation to the major axis of the axoneme (Farina et al. 1986). The PFR is composed of a large number of proteins, most of which have not been yet characterized. However, two major proteins that have been characterized in some detail are known as PFR 1 and 2, with molecular weights of 73 and 79 kDa, respectively. These proteins are highly antigenic and constitute targets for vaccine and diagnostic kit development. The available evidence indicates that the PFR is an essential structure for the parasite survival (Bastin & Gull 1999). The PFR is a good example of a complex structure well characterized in its morphology but where there is still no clear relationship between the structure and its composition despite the efforts made by several groups, including the use of a proteomics (Broadhead et al. 2006). The nucleus The initial observation showed a nucleus enveloped by typical membranes with pores (Fig. 13), a condensed chromatin dispersed throughout the nucleoplasm and a typical nucleolus found in the epimastigote (Fig. 14), but not in the amastigote or trypomastigote forms (De Souza & Meyer 1974, Elias et al. 2001). It was also shown that the nuclear membrane remains intact during the entire division process, with the appearance of intranuclear microtubules, dispersion of the chromatin and the appearance of dense plates whose number varies according to the trypanosomatid species (Solari 1995). Still, there is no evidence that these plates correspond to chromosomes that have been detected using a biochemical approach. The fibrillar structures (sub-pellicular microtubules) A fine cytoplasmic striation arranged in parallel conformation was observed in the first EM images of whole epimastigote forms (Fig. 2,3). These striations were mainly seen at the periphery of the protozoan, and treatment of the cell with trypsin or prolonged osmium fixation destroyed them. These striations were later seen in the first thin sections of cells fixed in osmium tetroxide, although the preservation was not very good. With the subsequent introduction of glutaraldehyde as the fixative (Sabatini et al. 1963), these structures were well preserved and designated as sub-pellicular microtubules. These microtubules are resistant to low temperature as well as to several drugs usually employed to disrupt the microtubules such as colchicines, colcemid, Taxol, etc. The analysis of replicas of quick frozen, freeze-fractured, deep-etched and rotary replicated cells, as developed by John Heuser and co-workers (Heuser 2001), clearly show that they are connected to each other through filaments (Fig. 15), as well as to the inner face of the plasma membrane and to the peripheral cisternae of the endoplasmic reticulum (Souto-Padron et al. 1984). Connections to components of the endocytic pathway are also observed. The three-dimensional organization of the microtubules can be seen in whole cells extracted with detergent, critical point dried and observed in a high resolution scanning electron microscopy (SEM). Negative staining of whole cells also revealed the sub-pellicular microtubules. Structures recognized after 1960 Glycosomes The examination of thin sections of several species of trypanosomatids revealed the presence of spherical structures surrounded by a unit membrane with a homogeneous matrix (Fig. 16). In some species, the organelle may display an elongated shape (Fig. 17). Initially, they were designated as microbodies, in analogy to similar structures described in mammalian cells, especially in hepatocytes. In mammalian cells, these structures were shown to contain catalase, an enzyme involved in the degradation of hydrogen peroxide formed in metabolic reactions and were thus named peroxisomes. Catalase was also found using a cytochemical approach in the microbodies of C. fasciculata (Muse & Roberts 1973) and in Leptomonas samueli (Souto-Padron et al. 1980). A major breakthrough in this field was the discovery that most of the glycolytic pathway takes place in this organelle (Opperdoes & Borst 1977). Because glycolysis generally takes place in the cytosol, the organelle was designated as a glycosome. Since catalase is observed in the glycosomes found in monogenetic trypanosomatids, but not in the digenetic ones, they are now considered to be a special type of peroxisome. EM cytochemistry aiming at the detection of basic proteins, as previously described in the kinetoplast section, showed intense labeling of the glycosomes (Souto-Padron & De Souza 1978). These observations anticipated the subsequent description, using a biochemical approach, that in contrast to what happens with the cytosolic glycolytic enzymes found in other eukaryotic cells the enzymes found in the glycosome show a higher isoelectric point. In addition to catalase, the peroxisomes of mammalian cells have more than 50 different enzymes involved in different metabolic pathways, such as peroxide metabolism, ß-oxidation of fatty acids, ether phospholipid synthesis, etc. Evidence has been obtained indicating that, in addition to the metabolic routes described above, other metabolic pathways such as carbon dioxide fixation (Opperdoes & Cotton 1982), purine salvage and pyrimidine biosynthesis de novo, fatty acid elongation, isoprenoid biosynthesis (review in Opperdoes 1987) also take place in the glycosomes of trypanosomatids, although they occur in the cytosol of other cells. The glycosome does not possess a genome. Therefore, all the proteins found within are encoded by nuclear genes, translated on free ribosomes and post-translationally imported into the organelle. The uptake of proteins into the glycosomes occurs within 5 min of protein synthesis and is due to the presence of several targeting signals (Shih et al. 1998). Vesicles and the endocytic pathway Two structures were initially identified as unique to T. cruzi and were later shown to be present in all members of subgenus Schyzotrypanum of genus Trypanosoma (T. dionisii, T. vespertiolinis, T. myoti among others). The first is the cytostome, which appears as an invagination of the plasma membrane and underlying microtubules that penetrates so deep into the cell that it may reach the nuclear region (Fig. 18). The opening of this structure has a diameter of about 0.3 µm and it is significantly smaller in the deeper portion. Subsequent studies showed that about 85% of the endocytic activity of T. cruzi epimastigotes takes place in the cytostome. Important information on the structural organization of the cytostome could be obtained using cryotechniques. Freeze-fracture initially showed that the membrane lining the cytostome corresponds to a highly specialized area (Martinez-Palomo et al. 1976, Pimenta et al. 1989) (Fig. 19). Additional important information was obtained using cryotechniques developed by Pinto da Silva and co-workers including label fracture and fracture-flip (Pinto da Silva et al. 1981, Anderson-Forsman & Pinto da Silva 1988). It was shown that the actual surface of the cytostome was rugous (Fig. 20) and it was heavily labeled with lectins (coupled to colloidal gold particles) (Fig. 21), thus indicating a high concentration of carbohydrate-containing molecules at this specialized portion of the cell surface (Pimenta et al. 1989). This latter observation was confirmed by high resolution SEM using backscattered electrons (Nakamura et al. 2005). The second unique structure to the Schyzotrypanum subgenus appears as a group of rounded or irregular structures mainly concentrated at the posterior region of the protozoan. They were first identified in thin sections and designated as multivesicular bodies since they present several inclusions in their matrix (Fig. 22). Subsequently, it was shown that the inclusions were surrounded by a single monolayer, rather than a unit membrane and contained lipids. Proteins were cytochemically detected in the matrix. Experiments where the protozoa were incubated in the presence of fluorescent- or colloidal gold-labeled proteins and LDL showed that these macromolecules are incorporated via typical pinocytic vesicles formed at the basis of the cytostome or in the flagellar pocket, which then fused to each other forming a long tubular structure. Immunocytochemical detection of cysteine proteinase, a major protease synthesized by the protozoan, showed that vesicles coming from the Golgi complex and containing this protein also fused with vesicles coming from the endocytic pathway. Subsequently, large organelles initially designated as multivesicular bodies that accumulate proteins ingested by endocytosis as well as those which are part of the secretory pathway are formed (Fig. 23). These observations led to the suggestion that this organelle was involved in the concentration of proteins ingested by the parasites. This organelle was designated as the reservosome. The reservosomes gradually disappear during transformation of epimastigotes into the infective trypomastigotes. The reservosome has also been characterized by the presence of proteases synthesized by the parasite, such as cysteine proteinase, as well as by the presence of small GTPases. The organelle was recently isolated and characterized biochemically (review in Cunha e Silva et al. 2006). Similar studies carried out with T. brucei and Leishmania have shown that all endocytic activity takes place in the flagellar pocket. In the case of T. brucei, these vesicles form a typical spherical lysosome while in Leishmania there is formation of a tubular structure (review in Field & Carrington 2004). Acidocalcisome Since the first observations of thin sections of T. cruzi as well as other trypanosomatids, a vacuolar structure containing an electron dense deposit was found and designated as polyphosphate granules or volutin granules. Only in 1994 was it shown that it was an organelle capable of transporting protons and calcium and was named an acidocalcisome. Since then, the organelle has been identified in all members of the Trypanosomatidae family and in many members of the Apicomplexa phylum (review in Docampo et al. 2005). Its morphology varies according to the protozoan species, the cultivation medium, etc. In some cells, the acidocalcisomes appear as rounded structures, with a diameter variable from 0.05 to 0.6 µm. It may also acquire an elongated shape and occupy about 2% of the cell volume. Its appearance depends on the methodology used to process the samples for EM. In conventional methods most of the dense content may disappear, leaving only a small electron dense dot associated with the membrane lining the organelle (Fig. 24). When cryofixation was used, a much better preservation of the acidocalcisome content was achieved. Indeed, a homogenous electron dense matrix was preserved when the cells were quick frozen, using the high pressure freezing technique and submitted to freeze-substitution (Fig. 25) (Miranda et al. 2000). The best way to visualize the acidocalcisomes is the observation of whole cells dried on the grid and observed using an electron microscope with an energy filter, as shown in Fig. 26. EM microanalysis has played a key role in the determination of the ionic composition of the acidocalcisome. Using X-ray mapping it was shown that they contain calcium, phosphorous, sodium, potassium and zinc (Fig. 27). In some trypanosomatids iron was also found (Miranda et al. 2004). The acidocalcisomes are involved in various functions including (i) storage of calcium, magnesium, sodium, potassium, zinc, iron, phosphorous compounds, especially inorganic pyrophosphate and polyphosphate, as determined by biochemical analysis and by X-ray microanalysis, (ii) pH homeostasis and (iii) osmoregulation, participating in close association with a contractile vacuole (review in Docampo et al. 2005). Contractile vacuole Although the presence of a contractive vacuole is very common in protozoa, there are few reports describing the presence of such a structure in trypanosomes. Following initial LM observations a structure resembling a contractile vacuole was characterized by EM (Fig. 28) to be formed by several tubules connected to a central vacuole located close to the flagellar pocket (Linder & Staehelin 1977, Attias et al. 1996). More recently aquaporin, a protein involved in water transport, was identified in epimastigotes of T. cruzi and showed to be located both in the acidocalcisomes and in the contractile vacuole (Fig. 29) (Montalvetti et al. 2004). These structures seem to be involved in the process of osmoregulation. It was shown that fusion of acidocalcisomes to the contractile vacuole occurs in a process mediated by cyclic AMP (review in Rohloff & Docampo 2008). The cell surface For many years, the predominant understanding was that the surface of the cells was smooth. With the use of cytochemical techniques designed to localize carbohydrate-containing molecules, such as the periodic acid-thiosemicarbazide-silver proteinate technique (ATSP) and the use of cationic dyes, especially ruthenium red, it became clear that all cells posses a structure presently known as the glycocalix or the surface structure. Such structure was also found in trypanosomatids. In this area the most interesting observation was the presence of a 15 nm thick surface coat in bloodstream forms of T. brucei (Vickerman 1969) (Fig. 31). Here, the initial EM observations made by Keith Vickerman opened a new and important area of research, including the identification of Variant Surface Antigens (VSGs) that make the surface coat and are responsible for the phenomenon of antigenic variation. However, other trypanosomes, including T. cruzi, do not show such a thick surface coat although a thin glycocalix could be demonstrated by EM cytochemistry using the periodic ATSP (review in De Souza 1989). Over the last 10 years several glycoconjugates (glycoproteins, glycolipids and lipophosphoglycans) have been identified and characterized in T. cruzi as well as in other trypanosomatids. One important feature is that most of these surface glycoconjugates are anchored to the protozoan plasma membrane via glycosylinositol anchors. Several of these molecules are involved in the interaction process of the protozoan with the host cell. Initially, the plasma membrane was considered as a unit membrane following the classical theory of Danieli and Davson and the unit membrane concept of Robertson. The introduction of the freeze-fracture technique allowed a better morphological characterization of the plasma membrane and the identification of special macro, micro and nanodomains of the cell membranes (review in De Souza 2007). In the case of T. cruzi it was shown that significant differences exist in the density and distribution of intramembranous particles, assumed to represent membrane integral proteins, according to the developmental stages. Additional differences were observed in the various regions of the membrane lining the cell body of the protozoan. For instance, in the epimastigote form it was clear that the flagellar membrane presents very few particles in contrast to the membrane lining the cell body (Fig. 19). The cytostome appeared as a particle-poor region delimited from the other portions of the membrane by a palisade-like row of particles. Specialized regions, where special arrays of particles exist, were observed in the region of attachment of the flagellum to the cell body membrane. Another highly specialized domain is the region of attachment of the flagellum to the cell body. Here, there are specializations, specially recognized in freeze-fracture replicas, on the fracture faces of the membrane lining the flagellum and the cell body (review in De Souza 2007). Perspectives EM will continue to provide important information on the structural organization of trypanosomes provided that technological advances continue. First, there are very few data on three-dimensional distribution of the various structures during the protozoan cell cycle. Certainly, important information can be obtained by three-dimensional reconstructions of the cell using both serial sections and EM tomography of epoxy sections. Second, more detailed information of some structures that can be isolated, such as the flagellum and the kinetoplast, can be obtained using cryoelectronic tomography. Third, EM analytical techniques, both using X-ray mapping and electron energy loss, will provide new information on the physiology of the organelles involved in ion transport. Fourth, immunocytochemistry of cryofixed cells will continue to add important information on the localization of new proteins discovered either by genomic or proteomic approaches. Fifth, atomic force microscopy is a powerful technique for high resolution analysis of some structures, especially those that are part of the cytoskeleton, as shown recently by Rocha et al. (2007) for structures existent on the flagellum. Sixth, high resolution SEM of the inner portion of the cells exposed either by mechanical rupture or by freeze-fracture presents a high potential to add new information concerning the relationships between the various structures. Seventh, it will be necessary to use EM to improve the resolution of the information obtained using confocal light immunofluorescence microscopy of GFP-labeled proteins. Finally, the various freeze-fracture techniques, especially in association with immunocytochemistry, will continue to be a powerful technique to analyze the structure of the cytoskeleton and various membranes. ACKNOWLEDGMENTS To several colleagues which have worked over the last 30 years in the author's laboratory, and David Straker for a critical reading of the manuscript. At the very beginning discussions with Hertha Meyer and Keith Porter opened new research areas. Later on, discussions with Adolfo Martinez Palomo, John Heuser, Pedro Pinto da Silva, Hans Muller, Vinci Mizuhira, Keith Vickerman and Jurgen Roth opened new perspectives for the study on the structural characterization of trypanosomes. REFERENCES
Copyright 2008 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc08055f2-6.jpg] [oc08055f1.jpg] [oc08055f7-9.jpg] [oc08055f18-23.jpg] [oc08055f10-12.jpg] [oc08055f30.jpg] [oc08055f25-27.jpg] [oc08055f28-29.jpg] [oc08055f16-17.jpg] [oc08055f13-15.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}