 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 103, No. 4, June, 2008, pp. 375-385 TNF/TNFR1 signaling up-regulates CCR5 expression by CD8+ T lymphocytes and promotes heart tissue damage during Trypanosoma cruzi infection: beneficial effects of TNF-α blockade Karina Kroll-PalharesI; Jaline Coutinho SilvérioI; Andrea Alice da SilvaII; Vladimir MichailowskyIII; Ana Paula MarinoI; Neide Maria SilvaIV; Cristiano Marcelo Espinola CarvalhoI; Luzia Maria de Oliveira PintoI; Ricardo Tostes GazzinelliIII; Joseli Lannes-VieiraI, + ILaboratório
de Auto-imunidade e Imuno-regulação (LAIIR), Instituto Oswaldo
Cruz-Fiocruz, Av. Brasil 4365, 21045-900 Rio de Janeiro, Brasil Financial support: CNPq (501561/2004-8, 301504/2005-9, 471518/2006-9), Faperj (E-26/151999/2000) Received: 17 March
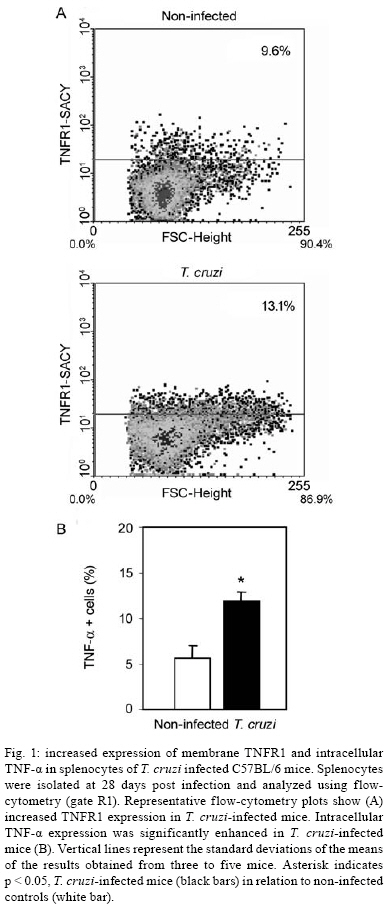
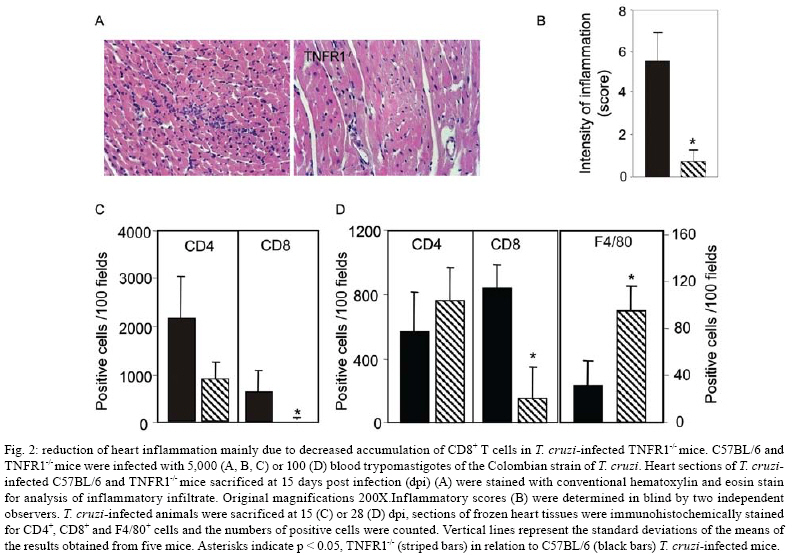
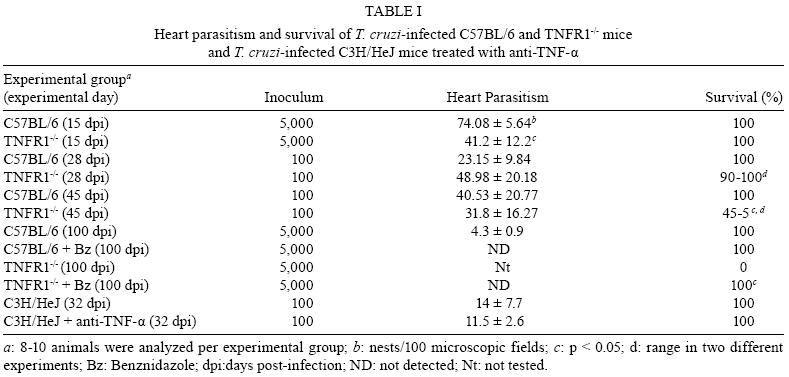
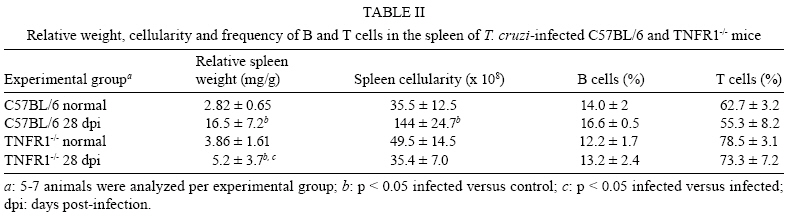
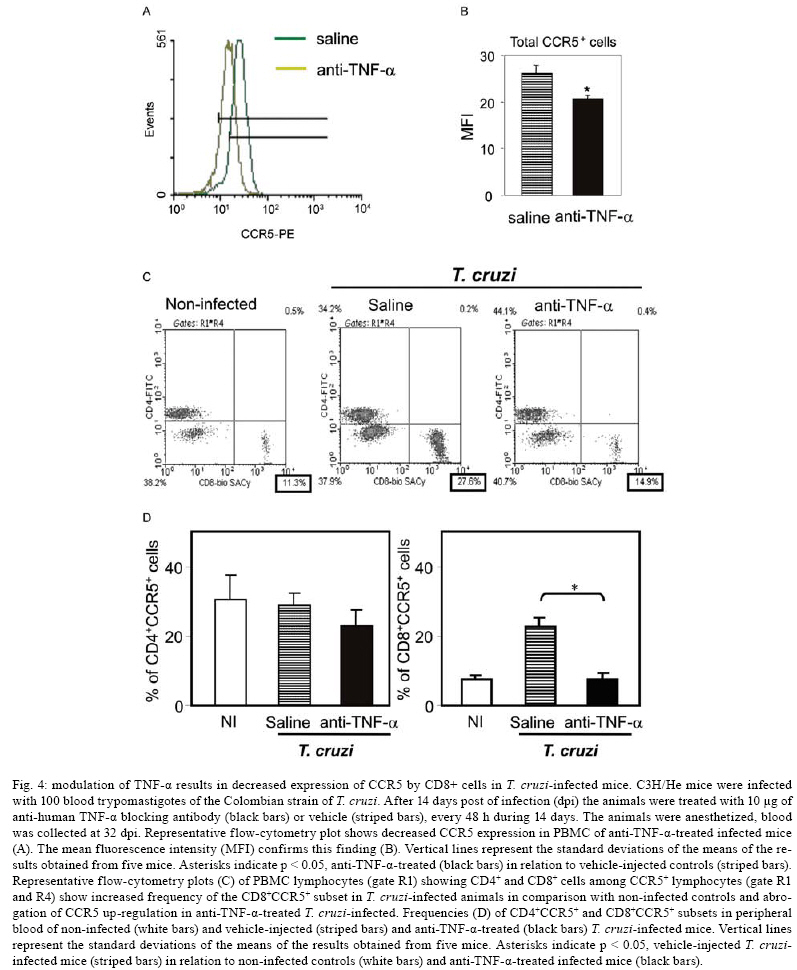
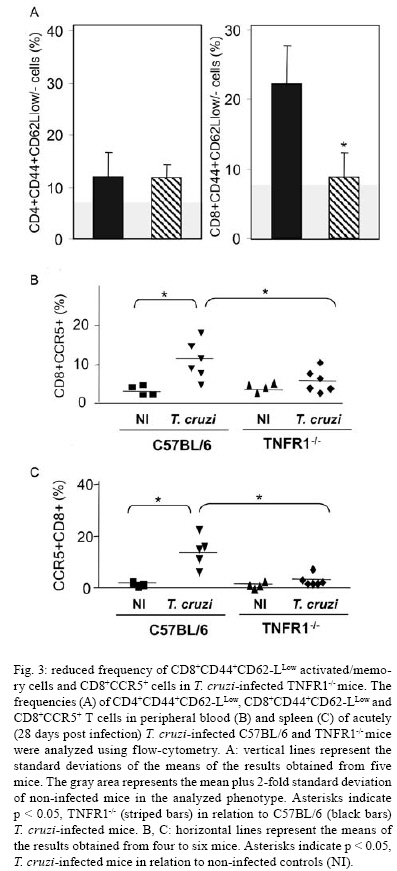
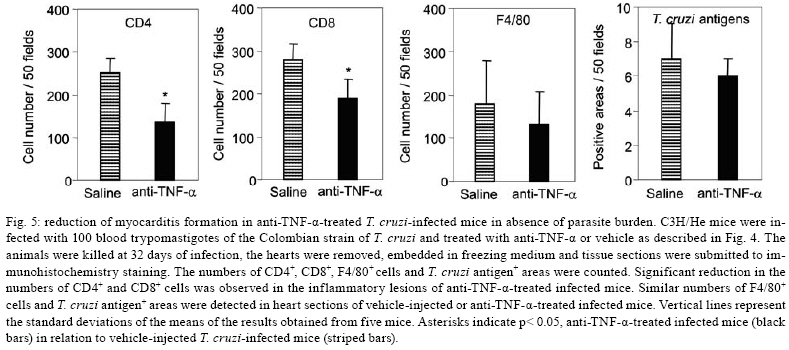
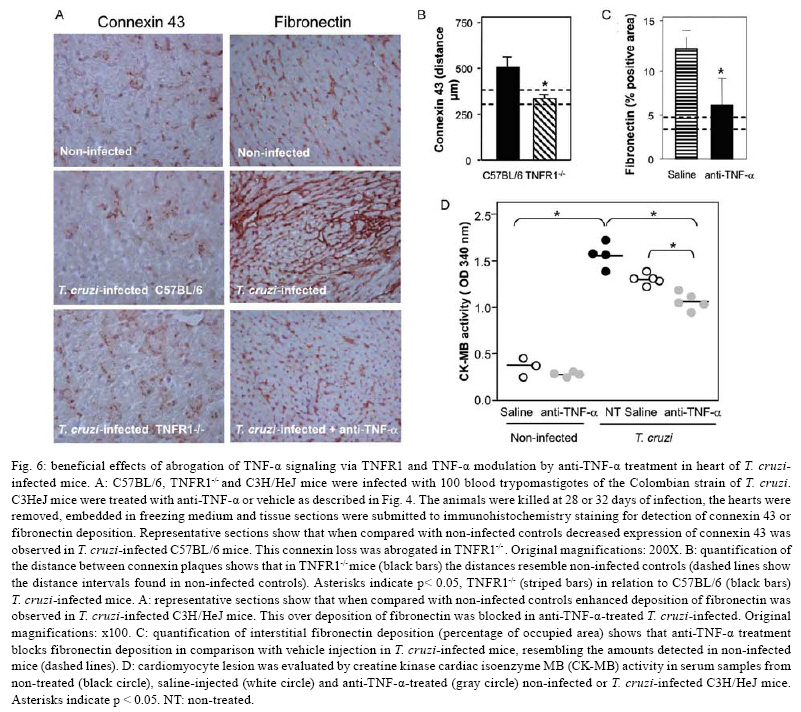
2008 Code Number: oc08065 ABSTRACT In Chagas disease, understanding how the immune response controls parasite growth but also leads to heart damage may provide insight into the design of new therapeutic strategies. Tumor necrosis factor-alpha (TNF-α) is important for resistance to acute Trypanosoma cruzi infection; however, in patients suffering from chronic T. cruzi infection, plasma TNF-α levels correlate with cardiomyopathy. Recent data suggest that CD8-enriched chagasic myocarditis formation involves CCR1/CCR5-mediated cell migration. Herein, the contribution of TNF-α, especially signaling through the receptor TNFR1/p55, to the pathophysiology of T. cruzi infection was evaluated with a focus on the development of myocarditis and heart dysfunction. Colombian strain-infected C57BL/6 mice had increased frequencies of TNFR1/p55+ and TNF-α+ splenocytes. Although TNFR1-/- mice exhibited reduced myocarditis in the absence of parasite burden, they succumbed to acute infection. Similar to C57BL/6 mice, Benznidazole-treated TNFR1-/- mice survived acute infection. In TNFR1-/- mice, reduced CD8-enriched myocarditis was associated with defective activation of CD44+CD62Llow/- and CCR5+ CD8+ lymphocytes. Also, anti-TNF-α treatment reduced the frequency of CD8+CCR5+ circulating cells and myocarditis, though parasite load was unaltered in infected C3H/HeJ mice. TNFR1-/- and anti-TNF-α-treated infected mice showed regular expression of connexin-43 and reduced fibronectin deposition, respectively. Furthermore, anti-TNF-α treatment resulted in lower levels of CK-MB, a cardiomyocyte lesion marker. Our results suggest that TNF/TNFR1 signaling promotes CD8-enriched myocarditis formation and heart tissue damage, implicating the TNF/TNFR1 signaling pathway as a potential therapeutic target for control of T. cruzi-elicited cardiomyopathy. Key words: heart disease - inflammation - Trypanosoma cruzi - CCR5 - TNFR1 - TNF-α Chagas disease, caused by the hemoflagellate protozoan Trypanosoma cruzi, afflicts 15-16 million people in South America with 75-90 million people exposed to infection (Coura 2007). Approximately 30% of patients develop a chronic disease typically characterized by myocarditis associated with prominent fibrotic scarring and organ dysfunction (Teixeira et al. 2006, Coura 2007). Although heart inflammatory cells contribute to control parasite growth, they are also involved in perpetuating heart disease (Freitas et al. 2005). CD8+ T cells predominate in myocardium from cardiac patients (Higuchi et al. 1997) and T. cruzi-infected mice (dos Santos et al. 2001). In experimental T. cruzi infection, the prevalence of CD8+ T cells in heart tissue has been linked with early production of the CC-chemokines CCL3/MIP-1α, CCL4/MIP-1β, and CCL5/RANTES as well as up-regulation of CCR5 on circulating leukocytes (Marino et al. 2004). Treatment with Met-RANTES, a partial CCR1/CCR5 antagonist, decreases cell infiltration in T. cruzi-infected murine hearts and has a beneficial effect on survival (Marino et al. 2004). In addition, infected CCR5-/- mice have deficient recruitment of T cells and macrophages to the heart (Hardison et al. 2006). These findings implicate CCR5 in the active development of T. cruzi-elicited myocarditis. Cytokines produced in the heart tissue of T. cruzi-infected individuals during the initial immune response can also influence the regulation of subsequent immune reactions (Brener & Gazzinelli 1997). TNF-α is detectable in inflamed hearts of chronic cardiomyophatic chagasic patients (D'Avila Reis et al. 1993), produced by T cells derived from endomyocardial biopsies (Abel et al. 2001). This suggests that TNF-α might be involved in the maintenance of chronic myocarditis. TNF-α signaling via TNFR1 (p55/60), but not TNFR2 (p75/80), plays a pivotal role in T. cruzi dissemination during acute experimental infection by controlling parasite uptake, nitric oxide (NO) as well as chemokine production, and cell mobilization to sites of infection (Aliberti et al. 2001). Importantly, TNF-α plasma levels directly correlate with the degree of heart dysfunction in chronic chagasic patients (Ferreira et al. 2003, Perez-Fuentes et al. 2003). In this context, the present study was undertaken to better understand the participation of TNF-α, especially signaling via TNFR1/p55, in the pathophysiology of T. cruzi infection, focusing on the mechanisms involved in cell activation, myocarditis development, and heart dysfunction. MATERIALS AND METHODS Animals - Five- to seven-week-old female C57BL/6 (H-2d) mice, B6.129-Tnfraf1a (p55/60)-deficient mice, and C3H/HeJ (H-2k) mice obtained from the animal facilities of the Oswaldo Cruz Foundation (Fiocruz, Rio de Janeiro) were maintained under standard conditions and manipulated according to the institutional guidelines for animal ethics and biosafety of Fiocruz (protocol #161/03). All B6.129-Tnfraf1a (p55)-deficient mice were genetically characterized using the primers and protocols described at The Jackson Laboratories home page (http://www.jax.org/). Experimental infection - Mice were infected intraperitoneally with 5,000, 1,000, or 100 blood trypomastigotes (bt) of the low virulence Colombian strain of T. cruzi isolated and maintained as previously described (dos Santos et al. 2001). Parasitemia was estimated using 5 µl of blood obtained from the tail vein, and employed as a parameter to establish acute and chronic phases (dos Santos et al. 2001). In some experiments, the animals were treated with a subcurative dose of Benznidazole (100 mg/kg/day) from days 10 to 17 after infection (Michailowsky et al. 2001). TNF blocking protocol - After 14 days of infection, C3H/HeJ (H-2k) mice were subcutaneously treated with injection grade saline (BioManguinhos-Fiocruz, Brazil) containing 10 µg of anti-human TNF-α blocking antibody (Remicade®, Infliximab), a gift from Schering-Plough of Brazil, at 48 h intervals over 14 days. Infliximab was previously shown to block in vivo TNF-α biological activities in murine models (Redlich et al. 2002). Antibodies and reagents - A polyclonal antibody recognizing T. cruzi antigens was produced in our laboratory (LAIIR). A polyclonal antibody recognizing mouse fibronectin was obtained from Gibco (USA). A polyclonal antibody recognizing connexin-43 was purchased from Sigma (USA). Purified anti-F4/80 antigen (clone F4/80) antibody was purchased from Caltag Laboratories (USA). Supernatants used for immunohistochemistry staining were produced in-house using anti-mouse CD8a (53-6.7) and anti-mouse CD4 (GK1.5) hybridomas. Biotin- and FITC-conjugated anti-mouse CD8a (53-6.7), PE- and biotin-conjugated anti-mouse CD4 (GK1.5), biotin-conjugated anti-CD62L (clone MEL-14), FITC-conjugated anti-CD44 (IM7.8.1), PE-conjugated anti-mouse TNF-α (MP6-XT22), PE-conjugated anti-mouse CCR5 (C34-3448), biotin-conjugated anti-ICAM-1 (CD54, clone 3E2), isotype control, and Cy-Chrome-streptavidin were purchased from BD PharMingen (USA). Biotinylated anti-rat immunoglobulin was purchased from Dako (Denmark). Biotinylated anti-rabbit immunoglobulin and peroxidase-streptavidin complex were purchased from Amersham (England). Appropriate controls were prepared by replacing primary antibodies with purified rat immunoglobulin or normal rabbit serum. Histopathology - Groups of 5-7 infected mice and 3-4 age-matched controls were killed under anesthesia at various time points post-infection. Hearts were removed and fixed in 4% buffered formalin. Sections were analyzed by light microscopy after paraffin-embedding and standard hematoxylin and eosin (H&E) staining. Tissue parasitism was scored by counting the total amastigote nests in each section (50-80 microscopic fields, 250x magnification). For the inflammatory infiltrate score, the number of inflammatory foci present in 50 microscopic fields was counted in each section (dos Santos et al. 2001). Immunohistochemistry (IHS) - The animals were killed under anesthesia at various time points post-infection. The hearts were removed, embedded in tissue freezing medium (Tissue Tek, Miles Laboratories, USA), and stored in liquid nitrogen. Serial 5 µm-thick serial cryostat sections were fixed in cold acetone and subjected to indirect immunoperoxidase staining or immunofluorescence, as previously described (dos Santos et al. 2001). Sections of spleen were used as positive controls for lymphocyte staining. Cells stained for CD4, CD8, and F4/80 were counted using light microscopy, with positive cells identified by a partial or complete rim of dark color outlining the cell membrane. Three sections were counted for each animal and the data are presented as average and standard deviation of each group of animals. The expression of connexin 43 and fibronectin in heart tissue sections was also evaluated using a digital area evaluation. Images were digitized using a color view XS digital video camera adapted to a Zeiss microscope. The images were analyzed using the AnaliSIS 3.2 Soft Image; two sections per heart and 25 fields per section were analyzed. The results are expressed as percentage of positive area per 25 mm2 of area in the heart or as the distance (µm) between gap junctions. Flow cytometric analysis - The animals were killed by blood removal under anesthesia. Suspensions of peripheral blood mononuclear cells (PBMC) were prepared using 0.3 ml of heparinized blood and performing Fycoll HypaqueTM(d = 1.077g/ ml) separation using a 3 ml column in a 5 ml tube (12x75mm, NUNC, USA). For intracellular cytokine staining, cells were isolated by individual samples and cells were incubated with 5 µg/ml of brefeldin A (Sigma, USA) at 37°C/ 4h. The cells recovered were washed, resuspended in PBS containing 2% fetal calf serum, and labeled as previously described (dos Santos et al. 2001). Controls for specific labeling were prepared using isotype matched controls. One-color labeled samples were prepared to set compensation values. Samples were acquired using a FACScalibur (Becton Dickinson, USA) gating on the PBMC and using a narrow forward-angle light scatter parameter to exclude dead cells from the analysis. At least 12,000 cells were acquired inside this gate. Fluorescence gates were cut in accordance to labeling controls, respecting curve inflexions. Cytometric analyses were performed using the program WinMDI Version 2.8. Statistical analysis - Data are expressed as arithmetic means ± standard error. Student's t test was used to analyze the statistical significance of the observed differences. The Kaplan-Meier method was used to compare the survival times of the studied groups. All statistical tests were performed with SPSS-8.0 (Michigan University, USA). Differences were considered statistically significant when p < 0.05. RESULTS T. cruzi infection increases expression of TNFR1 and TNF-α - When C57BL/6 mice were infected with 100 bt of the low virulence Colombian strain, surface TNFR1 and intracellular TNF-α were up-regulated in splenocytes at 28 days post-infection (dpi) (Figs 1a, 2b). TNF-α levels in serum and TNF-α mRNA in the heart tissue were increased during acute infection (28 dpi) and remained elevated throughout the chronic stage (120 dpi), as previously described (Starobinas et al. 1991, dos Santos et al. 2001). TNFR1 and TNF-α participate but are not critical for control of T. cruzi growth - TNFR1-/- mice infected with 5,000 or 100 bt exhibited increased parasitemia (e.g., when animals received 100 bt, 4.7 ± 2.4 x1 04 bt/ ml in C57BL/6 versus 19.2 ± 12.5 x 104 bt/ ml in TNFR1-/- at 28 dpi) and died from 25 to 50 dpi or from 28 to 100 dpi, respectively. C57BL/6 mice survived the acute infection independent of the number of parasites injected (Table I). Although cardiac parasitism was decreased at 15 dpi in TNFR1-/- mice infected with 5,000 bt, when TNFR1-/- mice received 100 bt a trend to increased parasitism was observed at 28 dpi. However, no parasite burden was observed in T. cruzi-infected TNFR1-/- mice when compared with C57BL/6 mice (Table I). Interestingly, in T. cruzi-infected C3H/HeJ mice treated with anti-TNF-α, parasitemia (3.4 ± 3.2 x 105 bt/ ml in non-treated versus 5.8 ± 3.8 x 105 bt/ ml in anti-TNF-α-treated mice at 32 dpi) and the numbers of heart parasite nests were similar to saline-injected controls (Table I). These data suggest that TNF-α and TNFR1 participate in but are not essential to the control of parasite growth. In fact, 100% of TNFR1-/- animals infected with 5,000 bt and treated with a subcurative dose of Benznidazole (100mg/ kg/ day) from day 10 to 17 dpi survived the acute infection and had no parasite nests in the cardiac tissue at 100 dpi (Table I). TNFR1 signaling is required for T. cruzi-elicited CD8-enriched myocarditis formation - The importance of TNF/TNFR1/p55 signaling in the development of myocarditis during acute T. cruzi infection was apparent in TNFR1-/- mice. In comparison with C57BL/6 mice, when TNFR1-/- mice were infected with 5,000 bt, myocarditis formation was hampered at 15 dpi (Figs 2a,b), in association with a decrease in the numbers of CD4+ and, mainly, CD8+ lymphocytes (Figs 2c). Furthermore, when TNFR1-/- mice infected with 5,000 bt were treated with a subcurative dose of Benznidazole, mice survived the acute phase and myocarditis formation was abrogated, in contrast to Benznidazole-treated wild-type mice which exhibited mild diffuse heart inflammation, reinforcing the role of TNFR1 signaling in myocarditis formation (data not shown). In the course of infection with 100 bt, hearts from C57BL/6 mice displayed a progressively increased percentage of infiltrating CD8+ T cells at 28 dpi, while few CD8+ cells infiltrated the hearts from infected TNFR1-/- mice (Figs 2d). In addition, T. cruzi-infected TNFR1-/- mice had a higher percentage of F4/80+ cells (macrophages) among their heart infiltrates than C57BL/6 mice (Figs 2d). The decreased myocarditis was not associated with re-compartmentalization of inflammatory cells to other muscle tissues as low numbers of CD4+, CD8+, and F4/80+ cells were found in the skeletal muscle of T. cruzi-infected TNFR1-/- mice compared to C57BL/6 mice (supplementary data). Interestingly, in infected TNFR1-/- mice, diminished myocarditis was associated with a decreased frequency of heart blood vessels expressing ICAM-1 (66.7 ± 4.5% in C57BL/6 versus 9.1 ± 6.8 % in TNFR1-/- at 28 dpi). Signaling via TNFR1 is required for complete CD8+ T-cell activation - To understand the molecular mechanisms hampering the formation of CD8-enriched myocarditis in T. cruzi-infected TNFR1-/- mice, we evaluated the numbers and activation phenotypes of mononuclear cells in the spleen and blood. T. cruzi infection of C57BL/6 mice resulted in a remarkable splenomegaly associated with enhanced cellularity; these effects were hindered in infected TNFR1-/- mice (Table II). Although the frequencies of total B and T cells were similar in both C57BL/6 and TNFR1-/- infected mice during the acute phase of infection (Table II), a significant accumulation of CD8+CD44+CD62Llow/- activated/memory lymphocytes was observed in the spleens of TNFR1-/- mice (4.56% ± 0.04% in C57BL/6 versus 11.67% ± 5.55% in TNFR1-/-), with a corresponding decrease of this cell population in peripheral blood (Figs 4a). This alteration was restricted to CD8+ T cells, as similar frequencies of CD4+CD44+CD62-Llow/- activated/memory T cells were seen in C57BL/6 and p55-deficient mice in both spleen (22.67% ± 2.04% in C57BL/6 versus 22.95% ± 10.69% in TNFR1-/-) and blood (Figs 3a). Since CCR5 is required to recruit T cells into cardiac tissue during T. cruzi infection (Marino et al. 2004, Hardison et al. 2006) and TNFR1-/- mice had defective cardiac infiltration and splenic retention of CD8+ cells, we studied the frequency of CD8+CCR5+ cells available for tissular recruitment in infected TNFR1-/- mice. During T. cruzi infection, an increased frequency of CD8+CCR5+ cells was detected in the blood (Figs 3b) and spleen (Figs 3c) of C57BL/6 mice, however these alterations were hampered in T. cruzi-infected TNFR1-/- mice. Altogether, these findings show the requirement for TNFR1 signaling for the acquisition of the full activation phenotype associated with migration potential in CD8+ lymphocytes during T. cruzi infection. Anti-TNF-α abrogates the up-regulation of CCR5 expression by CD8+ cells and myocarditis development during T. cruzi infection - To test the participation of TNF-α in controlling the expression of CCR5, particularly in CD4+ and CD8+ blood lymphocytes, anti-TNF-α blocking antibody (Infliximab) was administered during acute infection of C3H/HeJ mice, a model in which the participation of CCR5+ cells in T. cruzi-induced myocarditis formation was described (Marino et al. 2004). During acute infection by T. cruzi, a substantial increase in the numbers of circulating leukocytes was observed (4,000 ± 1,340 x 103/mm3 in non-infected versus 17,760 ± 2,478 x 103/mm3 in T. cruzi-infected mice). Treatment with anti-TNF-α did not alter the leukocytosis (13,100 ± 2,673 x103/mm3 in saline-injected versus 15,650 ± 2,800 x 103/mm3 in anti-TNF-α-treated), showing that this treatment did not exacerbate the aberrant leukocyte activation. Interestingly, the up-regulation of CCR5, measured as mean fluorescence intensity, by circulating leukocytes in T. cruzi-infected mice was prevented when the animals were treated with anti-TNF-α every 48 h from 14 to 32 dpi (Figs 4a,b). The abrogation of CCR5 up-regulation was mainly observed in CD8+ T cells (Figs 4c) and the frequency of CD8+CCR5+ T cells in anti-TNF-α-treated infected mice resembled that found in non-infected controls (Figs 4d). We next tested the effect of TNF-α blocking in myocarditis formation. A significant reduction in the numbers of CD4+ and CD8+ T cells was observed in the cardiac tissue of animals that received anti-TNF-α. In addition, the numbers of macrophages and areas positive for parasite antigens were similar in both groups (Figs 5). These results indicate that TNF-α is essential for CCR5 up-regulation involved in recruitment of lymphocytes to the heart during T. cruzi infection, and reinforce the hypothesis that massive heart inflammation is unrelated to the control of parasite growth. TNF/ TNFR1 signaling is involved in cardiac tissue damage during T. cruzi infection - TNF-α is potentially harmful to heart tissue in chronic Chagas disease (Ferreira et al. 2003, Perez-Fuentes et al. 2003). Thus, the functional integrity of myocardium was checked in TNFR1-/- and anti-TNF-α-treated infected mice through IHS using markers that are altered during T. cruzi infection, the gap junction marker connexin 43 (Campos-de-Carvalho et al. 1992) and fibronectin deposition (Marino et al. 2004). In non-infected controls, the expression of connexin 43 was regular and organized. However in T. cruzi-infected C57BL/6 mice, the expression of connexin 43 was reduced, disorganized, and more scattered, as characterized by increased distance between connexin spots. In contrast, in T. cruzi-infected TNFR1-/- mice, the expression of connexin 43 resembled that of non-infected controls (Figs 6a). Moreover, the distance between gap junctions in T. cruzi-infected TNFR1/p55-deficient mice was similar to non-infected mice, contrasting with T. cruzi-infected C57BL/6 mice (Figs 6b). When compared to non-infected controls, a remarkable increase in deposition of the extracellular matrix component fibronectin was detected in T. cruzi-infected C3H/HeJ mice. However, in anti-TNF-α-treated infected mice, the expression of fibronectin was significantly reduced and similar to non-infected controls (Figs 6a,b). More importantly, inspection of CK-MB levels, one of the markers of myocardial injury (de Souza et al. 2000), revealed that anti-TNF-α-treated mice had lower CK-MB levels compared to non-treated or saline-injected T. cruzi-infected C3H/HeJ mice (Figs 6d). Altogether, these results demonstrate that the TNF/TNFR1 signaling pathway is involved in heart tissue damage characterized by alterations in fibronectin deposition, electric conductivity, and cardiomyocyte injury during T. cruzi infection. DISCUSSION A collection of data implicates TNF-α in the progression of chagasic cardiomyopathy, yet the contribution of this cytokine to the pathophysiology of T. cruzi infection has not been elucidated. In the present study, we provide evidence that TNF-α, signaling through TNFR1/p55, is an inducer of cell activation, myocarditis development, and heart tissue damage during experimental T. cruzi infection. We observed an increase in TNFR1-bearing and TNF-α-expressing cells during infection of C57BL/6 mice with the low virulence Colombian strain of T. cruzi. TNF-α production was previously shown to be enhanced in T. cruzi-infected mice in association with susceptibility (Starobinas et al. 1991), and in chronically infected patients in association with heart dysfunction (Ferreira et al. 2003, Perez-Fuentes et al. 2003). T. cruzi-derived molecules, including glycosylphosphaditidylinositol-anchored mucins, have been shown to induce TNF-α production (Ropert et al. 2002). TNF-α synthesis can also be controlled by different intrinsic stimuli such as hormones, NO, cytokines, and other inflammatory mediators (Aggarwal et al. 2002). However, since several of these molecules are up-regulated in T. cruzi infection (Brener & Gazzinelli, 1997, Silva et al. 2003), the molecular circuit leading to TNF-α overproduction in this disease remains unclear. Previous studies using TNFR1-FcIgG3 transgenic and TNFR1-/- mice showed that TNF-α is required for the control of parasite burden during T. cruzi infection (Santos-Lima et al. 1997, Aliberti et al. 2001). TNFR1-/- mice treated from 10-17 dpi with a subcurative dose of Benznidazole survived the acute infection and exhibited reduced heart inflammation during the chronic phase, contrasting with IFN-γ-/- and IL12-/- mice, which succumbed shortly after treatment cessation with intense parasitemia, parasitism, and inflammatory processes (Michailowsky et al. 2001). Therefore, though important, TNF-α is not crucial for the control of parasite dissemination. Our results show that TNFR1 is crucially involved in the development of T. cruzi splenomegaly. Splenomegaly induced by T. cruzi infection has been linked to aberrant B and T cell activation (Minoprio 2001), especially CD8+ T-cell dysregulation (dos Santos et al. 2001, Tzelepis et al. 2007). Additionally, immunological abnormalities were linked to T. cruzi persistence and toxic lesions (DosReis et al. 2005). Thus, our data suggest that TNF/TNFR1 signaling might play a detrimental role during T. cruzi infection by contributing to the development of immunological abnormalities, mainly involving CD8+ T cell activation, and heart injury. We have previously proposed that CD8-enriched T. cruzi-elicited myocarditis formation is dependent on the differential activation of the CD8+ T-cell population after infection (dos Santos et al. 2001). In support of this model, the paucity of CD8+ T cells in cardiac tissues of TNFR1-/- infected mice correlated with a decreased frequency of activated/memory CD44+CD62LlowCD8+ cells in the blood and retention of this cell subset in the spleen. These data indicate that TNF/TNFR1 signaling is not essential for the full differentiation of this T-cell subset, as indicated by the expression of the activation/ memory markers CD44 and CD62L, but somehow controls CD8+ T-cell compartmentalization. CCR5 up-regulation, particularly by CD8+ T cells, occurs during T. cruzi infection (Marino et al. 2004). However, when TNF signaling was disrupted in TNFR1-/- mice or by anti-TNF-α treatment of T. cruzi-infected mice, a decrease in the frequency of CD8+CCR5+ T lymphocytes was observed in the peripheral blood and spleen, and was associated with reduced heart inflammation. Thus, our results indicate that TNF/TNFR1 signaling is a pre-requisite for CCR5 expression by CD8+ T cells, promoting CD8-enriched myocarditis formation. Nonetheless, future studies are required to elucidate the correlation between TNF-α and CCR5 modulation, including the involvement of intracellular signaling and transcriptional factors such as AP-1 and NF-kB, both of which are parts of the TNFR1 signaling pathway (Baud & Karin 2001). T cells, mostly CD8+, present in the myocardium of T. cruzi-infected mice express CCR5 (Marino et al. 2004) in an environment enriched in TNF-α and CC-chemokines (dos Santos et al. 2001, Marino et al. 2004). CCR5 up-regulation by CD8+ T cells allows their attraction to sites where the cognate chemokines are produced (Castellino et al. 2006). Partial blockade of CCR5 in T. cruzi-infected mice decreased myocarditis without hampering the control of parasite growth (Marino et al. 2004). In the present study, we demonstrated that interfering with the biological effects of TNF-α using Infliximab, an antibody that blocks soluble and membrane-bound TNF-α (Wong et al. 2008), led to CCR5 down-modulation and ameliorated heart inflammation without interfering with parasite control. On the other hand, infection of CCR5-/- mice results in higher parasite burden, mortality, and impaired macrophage and T cell influx into the cardiac tissue (Hardison et al. 2006). Therefore, we believe that the total absence of either TNFR1 or CCR5 signaling impairs the entrance of parasite-controlling cells in the heart, while the partial reduction of TNF-α and CCR5 signaling promotes beneficial effects for cardiomyopathy. Furthermore, the decreased recruitment of cells to cardiac tissue in TNFR1-/- infected mice was associated with diminished ICAM-1 expression by heart endothelial cells. Thus, it seems that the TNF/TNFR1 signaling pathway plays a crucial role in inflammatory cell extravasation to cardiac tissue by controlling the expression of CCR5 and ICAM-1, molecules involved in the establishment of T. cruzi-elicited myocarditis (Marino et al. 2004, Michailowsky et al. 2004). The reduced myocarditis observed in TNFR1-/- and anti-TNF-α-treated infected mice compared to the appropriate controls was associated with diminished tissue damage, indicated by preserved myocardial cell connectivity and decreased fibronectin deposition, resembling non-infected controls. The expression of connexin 43, a component of cardiac gap junctions leading to myocardium electric connectivity (Delmar et al. 2004), is profoundly altered in cardiomyocytes infected in vitro with T. cruzi (Campos-de-Carvalho et al. 1992). However, it remains to be clarified whether this is a direct effect of the parasite. Herein, we observed that abrogation of TNF/TNFR1 signaling preserves connexin 43 patterns in the cardiac tissue of T. cruzi-infected mice. TNF-α has important roles in the depression of heart function by down-regulating connexin 43 (Fernandez-Cobo et al. 1999). In this context, our data suggest that during T. cruzi infection TNF-α might play a role in reducing connexin 43 expression. Enhanced deposition of extracellular matrix components in the inflamed cardiac tissue has been described in T. cruzi infection (Andrade et al. 1991, Marino et al. 2004). We observed that heart fibronectin deposition was diminished in anti-TNF-α-treated infected mice, in parallel with a reduction in inflammation. Interestingly, in Verapamil-treated T. cruzi-infected mice, in situ modulation of TNF-α and IL-1β correlates with reductions in heart fibrosis and inflammation (Huang et al. 1999). These findings reinforce the hypothesis that TNF-α blockade might exert a beneficial effect during T. cruzi infection by modulating heart fibrosis or fibrogenic inflammation. Lastly, we found that during T. cruzi infection, anti-TNF-α treatment partially hampered the development of cardiomyocyte lesions, characterized by the release of CK-MB (de Souza et al. 2000), reinforcing the role of TNF-α as a contributor to cardiac injury. Recently, TNF-α blockade has been shown to decrease necrotic areas in the spleen during acute T. cruzi infection (Andrade et al. 2008). Conversely, treatment with Etanercept (soluble human TNFR2/p75 that binds TNF-α and lymphotoxin α) aggravated chronic chagasic cardiomyopathy in hamsters, leading to the claim that the absence of TNF-α signaling may be deleterious to the failing heart in Chagas disease cardiomyopathy (Bilate et al. 2007). This apparent paradox emphasizes the need for an understanding of the modes of action and the limiting factors of emerging novel therapeutic tools that target TNF-α (Wong et al. 2008). Furthermore, TNF-α is involved in cytoprotective signals that prevent and/or delay the development of cardiomyocyte apoptosis (Kurrelmeyer et al. 2000). However, it is clear that prolonged exposures to high levels of TNF-α have deleterious effects on cardiac function (Sarzi-Puttini et al. 2005). In fact, chronic chagasic patients with left ventricular dysfunction had 2-fold higher TNF-α levels than patients without ventricular dysfunction (Ferreira et al. 2003). In addition, cumulative and complementary effects of TNF-α and NO levels correlate with adverse prognosis in chagasic patients (Perez-Fuentes et al. 2003), suggesting that susceptibility to severe chagasic cardiomyopathy is a multi-factorial process in which TNF-α plays a protagonist role. Thus, the biological roles played by TNF-α (and other inflammatory cytokines such as lymphotoxin ) in T. cruzi-elicited cardiomyopathy deserve to be further explored for the development of rational therapeutic strategies. In this context, our results suggest that the detrimental effects of TNF-α in T. cruzi infection might be determined by the degree and duration of TNF-α production. Therefore, modulation (but not complete abrogation) of TNF-α production or its biological action might have a beneficial effect by keeping parasite dissemination under control, while impairing the development of harmful myocarditis and cardiomyocyte lesions during T. cruzi infection. REFERENCES
Copyright 2008 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc08065f5.jpg] [oc08065f4.jpg] [oc08065f6.jpg] [oc08065t1.jpg] [oc08065f1.jpg] [oc08065t2.jpg] [oc08065f2.jpg] [oc08065f3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}