 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 103, No. 5, August 2008, pp. 468-471 Leptoconops nosopheris sp. n. (Diptera: Ceratopogonidae) and Paleotrypanosoma burmanicus gen. n., sp. n. (Kinetoplastida: Trypanosomatidae), a biting midge - trypanosome vector association from the Early Cretaceous George Poinar Jr. Department of Zoology,
Oregon State University, Corvallis, OR 97331, USA Received 20 February
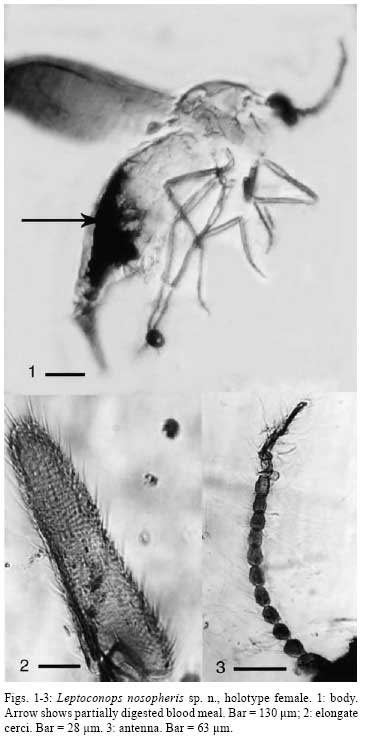
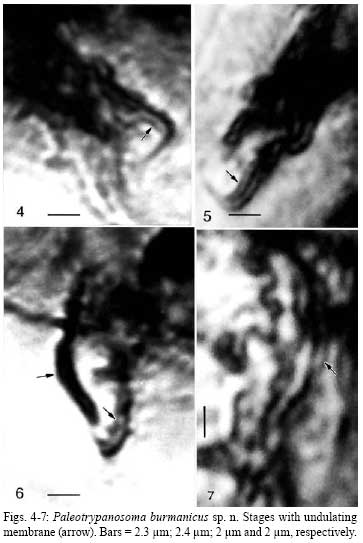
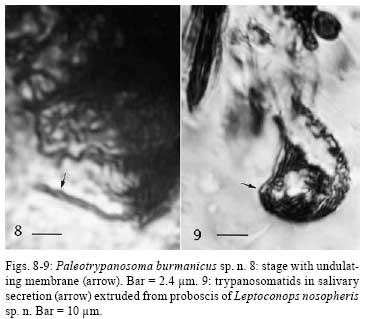
2008 Code Number: oc08082 ABSTRACT Leptoconops nosopheris sp. n. (Diptera: Ceratopogonidae) is described from a blood-filled female biting midge in Early Cretaceous Burmese amber. The new species is characterized by a very elongate terminal flagellomere, elongate cerci, and an indistinct spur on the metatibia. This biting midge contained digenetic trypanosomes (Kinetoplastida: Trypanosomatidae) in its alimentary tract and salivary glands. These trypanosomes are described as Paleotrypanosoma burmanicus gen. n., sp. n., which represents the first fossil record of a Trypanosoma generic lineage. Key words: Cretaceous - biting midge - trypanosomatids - fossil The fossil record of trypanosomatids is limited to Paleoleishmania proterus Poinar and Poinar (2004) vectored by an Early Cretaceous sand fly in Burmese amber and Trypanosoma antiquus Poinar vectored by a triatomid bug in Tertiary Dominican amber (Poinar 2005). The present study reports a novel association between a new species of biting midge belonging to the genus Leptoconops (Diptera: Ceratopogonidae) and a new species of associated trypanosome in Cretaceous Burmese amber. The trypanosomes were located in the gut, salivary glands and salivary secretions of the biting midge. The amber was derived from a mine first excavated in 2001, in the Hukawng Valley, Southwest of Maingkhwan in the state of Kachin (26º20´N, 96º36´E) in Burma, (Myanmar). This new amber site, known as the Noije Bum 2001 Summit Site, was assigned to the Upper Albian of the Early Cretaceous on the basis of paleontological evidence (Cruickshank & Ko 2003), placing the age at 97 to 110 m.y.a. Nuclear magnetic resonance spectra and the presence of araucaroid wood fibers in amber samples from the Noije Bum 2001 Summit site indicate an araucarian (possibly Agathis) tree source for the amber (Poinar et al. 2007). MATERIALS AND METHODS The amber piece containing the fossil biting midge with its associated trypanosomes was first shaped with a Lortone Model FS Lapidary Trim saw, then sanded on a Highland Park 1009 sander and finally polished on a Black and Decker 5 inch model 7900 bench grinder, during which time the specimen was observed with a Nikon SMZ-10 R stereoscopic microscope. The final piece is roughly rectangular in outline, measuring 12 mm long by 7 mm wide and 1 mm in depth. The flagellates were observed and photographed with a Nikon Optiphot compound microscope with magnifications up to 1,050X. The insect's tissues had partially cleared (Fig.1), thus allowing an unobstructed view of the alimentary tract, body cavity and salivary glands. The flagellates were preserved by natural fixatives in the original plant resin, which enter the cells soon after contact is made with the deposit. The method of insect preservation in resins is discussed by Poinar and Hess (1982, 1985). With this natural fixation process, most flagellates appear as opaque or translucent outlines under transmitted light. When visible, nuclei appear as large dark objects, while kinetoplasts are represented as small dark objects. The undulating membranes can be detected as wavy, transparent zones located along the side of the flagellates. It is difficult to photograph such small microbes through both the amber matrix and insect's body wall and the images are not as clear as extant forms are on a microscope slide. The images of the trypanosomes were enhanced in Adobe Photoshop by manipulating the brightness, contrast and magnification. Leptoconops nosopheris sp. n. (Fig.1-3) Female - (Fig.1) Body length 1.2 mm; antenna (Fig.3) with 12 flagellomeres; length antenna, 322 µm; flagellomeres nearly spherical in shape, slightly longer than wide; flagellomere 12 narrow, 82 µm long; 2.9 times as long as flagellomere 10 (flagellomere 11 is slightly compressed) (Fig.3); proboscis relatively long; palp four-segmented with third palpomere wider than others; eye separation not visible; hind tibia with indistinct apical spur; claws small, simple; wing length, 626 µm; costa slightly prolonged beyond vein R3; membrane covered with distinct microtrichia; macrotrichia absent; CR = 0.20; abdomen slender; cerci very long, 206 µm, 4.1 times as long as wide at the widest portion (Fig.2). Male - unknown. Type locality - Amber mine in the Hukawng Valley, Southwest of Maingkhwan in the state of Kachin (26º20´N, 96º36´E), Northern Myanmar (Burma). Type material - Holotype female in Burmese amber piece, deposited in the Poinar amber collection (code B-D-47b) maintained at Oregon State University, USA. Etymology - Noso is from the Greek for disease, sickness and pheris is from the Greek phero, to carry or bear. The biting midge had the remains of a blood meal associated with masses of trypanosomes in its midgut. The peritrophic membrane was no longer evident and most of the trypanosomes were distributed along the outer edge of the remaining brownish-yellow residue. No trypanosomes could be seen in the hemocoel, but they were quite abundant in the salivary glands and salivary ducts and some had been expelled from the tip of the insect's proboscis in a droplet of salivary secretion. All of the forms were relatively slender, which may have been due to shrinkage during the fossilization process in the amber. While it was difficult to see the position of the kinetoplast in relation to the nucleus in most specimens, some had the orientation of the nuclei and kinetoplasts characteristic of trypomastigotes and epimastigotes, as well as undulating membranes along the border of their bodies. Paleotrypanosoma gen. n. General diagnosis: Trypanosomatine: Trypanosomatidae. Paleotrypanosoma gen. n. is established for fossil members of the generic lineage Trypanosoma associated with extinct species of biting insects. The remarkable diagnostic characters of Paleotrypanosoma gen. n. are its age, presence of slender stages, absence of amastigotes and occurrence in an extinct species of biting midge. Paleotrypanosoma burmanicus sp. n. (Fig.4-7, 8-9) The description is based on epimastigotes and trypomastigotes located along the edge of the blood residue in the gut (Fig.1) and in the salivary droplet issuing from the proboscis of the biting midge (Fig. 9). Trypanosomes (n = 15) - Average length = 12 µm, range = 9-15 µm; average width = 1.8 µm, range = 1.3-2.0 µm; average length of flagellum = 6 µm, range = 5-7 µm. Some variation was noted regarding the positions of the kinetoplast and nuclei. Trypanosomes with the kinetoplast in the posterior portion of the body were regarded as trypomastigotes while those with the kinetoplast anterior to the nucleus were considered epimastigotes. Type material - Trypomastigote in the gut of L. nosopheris s. n. in Burmese amber deposited in the Poinar amber collection (code B-D-47b) maintained at Oregon State University, USA. Type host: L. nosopheris sp. n. (Diptera: Ceratopogonidae) in Burmese amber; vertebrate host unknown. Type locality - Amber mine in the Hukawng Valley, southwest of Maingkhwan in the state of Kachin (26º20´N, 96º36´E), Northern Myanmar (Burma). Etymology - The specific name of this protozoan species refers to country where is located its type locality, Burma. DISCUSSION Females of the genus Leptoconops feed on vertebrate blood from reptiles, birds and mammals (Auezova et al. 1990, Mullens et al. 1997, Mullen 2002, Szadziewski & Poinar 2005). The genus extends back some 130 million years with species found in Lebanese amber (Poinar & Milki 2001). There are 134 extant and, with the present specimen, 14 fossil species, five of which occur in Burmese amber (Szadziewski & Poinar 2005). This is the first species of Leptoconops, extant or extinct, associated with trypanosomes. Both digenetic and monogenetic trypanosomes have been reported from extant biting midges (Sharp 1928, Kremer et al. 1961, Baker 1976, Podlipaev et al. 2004b, Svobodová et al. 2007) and a monogenetic species is known from a biting midge in Burmese amber (Poinar & Poinar 2005). One extant trypanosome isolated from a biting midge was closely related to a strain isolated from Egyptian rats and could have been transferred to the vertebrate by the insect during blood feeding (Podlipaev et al. 2004a). Since biting midges take blood from all classes of vertebrates, it is not possible to determine the host of P. burmanicus sp. n. Many workers implicate biting midges (together with black flies and mosquitoes) as carriers of bird trypanosomes with transmission through contamination or ingestion of the vector (posterior station transmission) (Bennett 1961, 1970, Desser et al. 1975, Baker 1976, Miltgen & Landau 1982, Votypka & Svobodová 2004). However metacyclic trypomastigotes were noted in the mouth parts and salivary glands of some of the vectors and inoculative experiments showed that these forms were infective (Bennett 1970, Baker 1976). Since epimastigotes and trypomastigotes were found in the alimentary tract, salivary glands and salivary secretions of L. nosopheris sp. n., P. burmanicus sp. n. is considered to be a digenetic trypanosome that is transmitted by the bite of the biting midge. The life cycle of P. burmanicus sp. n. is probably similar to that of Trypanosoma b. brucei in the tsetse fly, Glossina m. morsitans. In the tsetse fly, the trypanosomes first multiply in the midgut, then migrate anteriorly to the foregut and finally into the salivary glands. Vertebrate-infective trypanosomes also occurred in salivary exudates which were extruded by the infected tsetse flies (Peacock et al. 2007). All studied biting midge- transmitted trypanosomes that infect birds are carried by species of the genus Culicoides Latreille (Fallis & Bennett 1961, Bennett 1961, 1970, Baker 1976, Miltgen & Landau 1982, Linley 1985). However it has been speculated that present day bird trypanosomes evolved from species that parasitized reptiles (Baker 1976), so it is possible that P. burmanicus sp. n. was parasitizing reptiles 100 million years ago. Support for a reptilian host are studies showing that several extant species of Leptoconops feed on lizards (Auezova et al. 1990, Mullens et al. 1997) and tortoises (Moravec & Orszagh 1998, Široký et al. 2007). There is evidence that vertebrate parasitism by trypanosomatids evolved multiple times in the past (Fernandes et al. 1993) and that the earliest trypanosomatid lineage was T. brucei. While this species is vectored by tsetse flies today, it is possible that in the Early Cretaceous, before tsetse flies evolved, precursors of the T. brucei or a now extinct lineage were being vectored by biting midges. It is possible that biting midges were the original ancestral vectors of the Trypanosoma lineage, since vertebrate feeding ceratopogonids extend back some 130 million years (Poinar & Milki 2001). Later, these flagellates could have been acquired by other biting Diptera, which acted as bridge vectors and transferred the parasites to new vertebrate hosts. ACKNOWLEDGMENTS To RL Jacobson, for providing references and discussions during the course of this study, and Roberta Poinar for comments on earlier versions of the manuscript. REFERENCES
Copyright 2008 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc08082f4-7.jpg] [oc08082f10.jpg] [oc08082f1-3.jpg] [oc08082f8-9.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}