 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
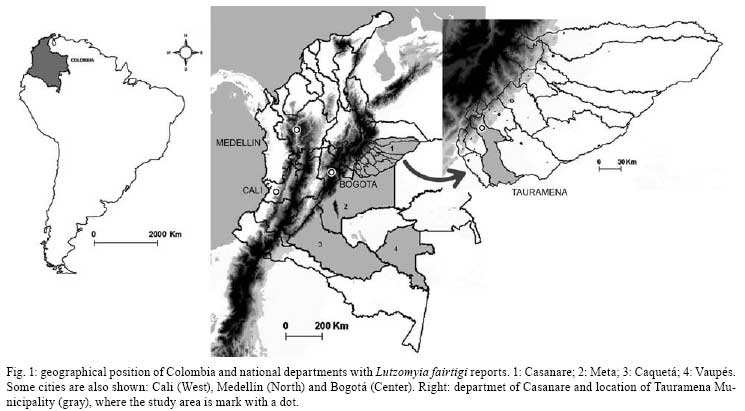
Memórias do Instituto Oswaldo Cruz, Vol. 103, No. 5, August 2008, pp. 477-482 Seasonality of Lutzomyia fairtigi (Diptera: Psychodidae: Phlebotominae), a species endemic to Eastern Colombia Jorge Alberto Molina*; Mario Iván Ortiz; Felipe Guhl Centro de Investigaciones
en Microbiologia y Parasitologia Tropical, Universidad de Los Andes, Departamento
de Ciencias Biológicas, Apartado Aéreo 4976, Bogotá, Colombia Received 8 March
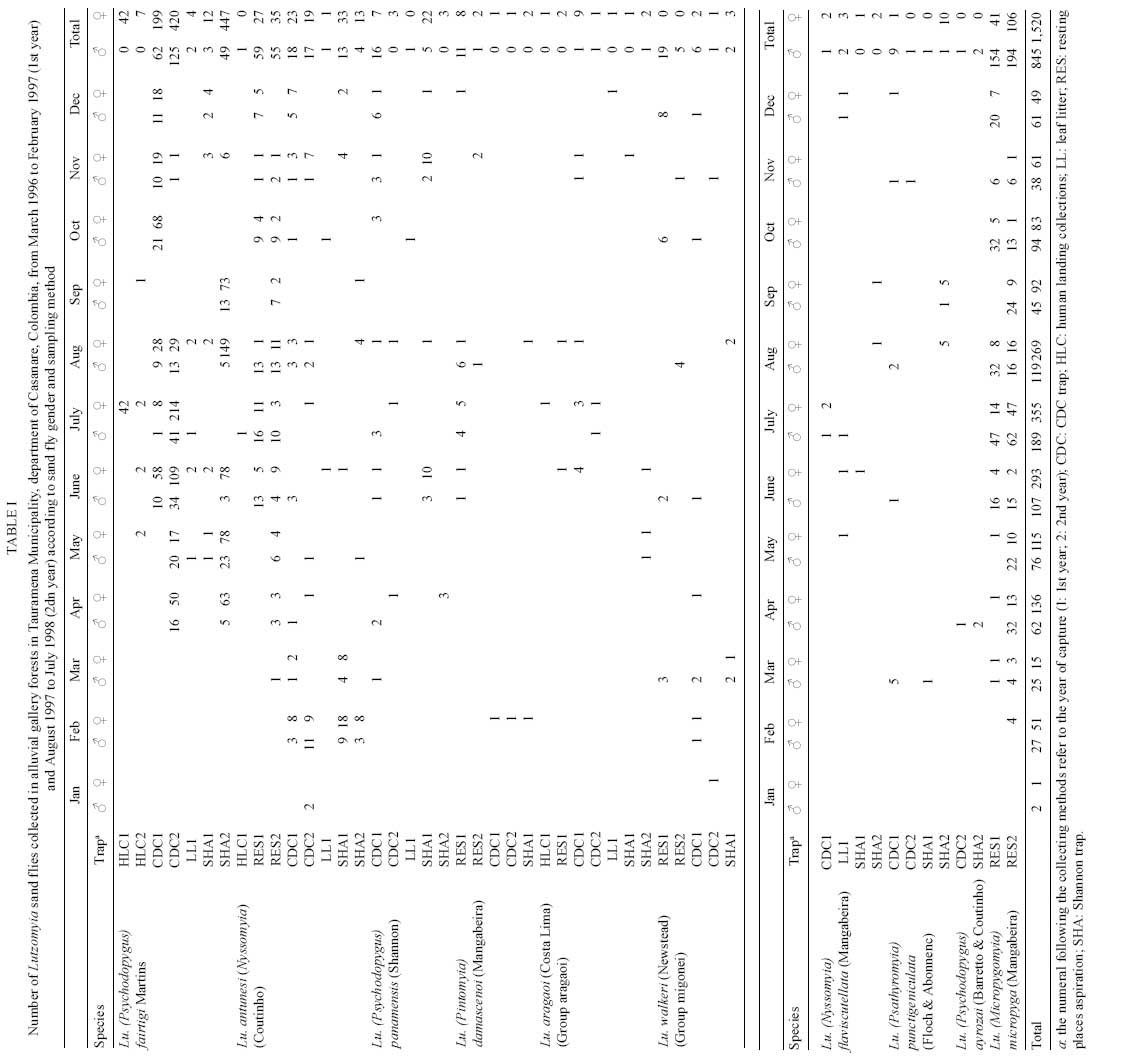
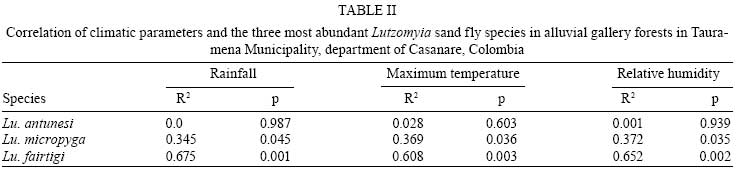
2008 Code Number: oc08084 ABSTRACT The bionomics of sand flies (Diptera: Phlebotominae) was studied monthly for two consecutive years in alluvial gallery forests in the department of Casanare, Northeastern Colombia. A total of 2,365 specimens and 10 species were captured using CDC light traps and Shannon traps, and from diurnal resting places, and human landing collections. Lutzomyia fairtigi Martins (55%), Lutzomyia micropyga (Mangabeira) (20.9%), and Lutzomyia antunesi (Coutinho) (13.5%) were the predominant species in the region. Lutzomyia flaviscutellata and Lutzomyia panamensis, potential vectors of Leishmania in Colombia and neighboring countries, were also collected, but in low numbers. Lu. fairtigi is an endemic species to Colombia, and minimal data are available on its biology and distribution. The present study provides additional information about Lu. fairtigi, such as the diurnal activity displayed by females on cloudy days, the greater density during the rainy season (April to October), marked anthropophilia, and the presence of flagellates in the midgut of one female. Key words: Phlebotominae - Lutzomyia fairtigi - Colombia - leishmaniasis - sand flies Phlebotomine sand flies are the vectors of leishmaniasis and phleboviruses, both of which cause important health problems and diseases in humans and domestic animals (Alexander 2000). In Colombia, 141 species of Phlebotomine sand flies have been reported (Bejarano 2007), and 27 of them are endemic to Colombia. Most of the endemic species have been reported with very little data, such as a first description of the species and preliminary geographic distribution. Consequently, additional important epidemiological information is lacking for most of these species. The only Psychodopygus species endemic to Colombia is Lutzomyia fairtigi Martins (Williams 1999). Lu. fairtigi was collected and described for the first time in forests close to Villavicencio (department of Meta, ME) using light and Shannon traps (Martins 1970). Bejarano (2007) showed that the geographical distribution of Lu. fairtigi is restricted to the eastern part of Colombia, in the departments of Caquetá, Casanare (CA), ME and Vaupés (Fig. 1). However, no information related to its epidemiological status has been reported (Young & Duncan 1994) with regard to natural Leishmania parasite infections and/or feeding habits. Leishmaniasis is an endemic disease in Colombia (except in San Andrés, Atlántico and Bogotá) (Corredor et al. 1990). In 2007, 5,771 cases were reported (97.7% cutaneous, 1.39% mucocutaneous, and 0.9% visceral) (SIVIGILA; www.ins.gov.co/nivel3.php?seccion=26&sub=45). The Leishmania species isolated from humans, animals, and sand flies were L. (Viannia) panamensis (53.8%), L. (Viannia) braziliensis (30.3%), L. (Leishmania) infantum (9.4%), L. (Viannia) guyanensis (2.6%), L. (Leishmania) amazonensis (1.8%), and L. (Leishmania) mexicana (0.8%) (Corredor et al. 1990). More recently, similar results were obtained from patients arriving in Bogotá from different regions of Colombia: L. panamensis (74.45%), L. braziliensis (15.33%), L. guyanensis (0.73%), L. mexicana (5.11%), and L. mexicana complex (3.65%) (Ovalle et al. 2006). In the departments of Caquetá, CA, ME and Vaupés, a total of 31,282 cases of leishmaniasis have been reported since 2000 (99.7% were cutaneous, 0.24% mucocutaneous, and 0.03% visceral) (SIVIGILA). The parasite species circulating in these four departments were L. panamensis, L. braziliensis, L. amazonensis, L. guyanensis, and L. mexicana (Corredor et al. 1990, Ovalle et al. 2006). In the same departments, 66 species of Lutzomyia were reported, six of which are vectors of Leishmania in Colombia (Bejarano 2007). To improve our knowledge of leishmaniasis transmission in CA, two consecutive years of entomological surveillance were carried out in this department. The main objectives of the survey were to determine monthly and annual trends in the species composition, monthly densities, and infection rates of sand flies. MATERIALS AND METHODS Study area - Sand flies were collected in Tauramena Municipality, CA (5°00'N, 72°42'W) in the Eastern Cordillera of the Colombian Andes (Fig. 1). The region is described as tropical premontane wet forest with some elements of tropical wet forest (IGAC 1988) located approximately 450 m above sea level. On average, the annual rainfall is 2,842 mm, mainly occurring between April and October. The annual temperatures range from a minimum of 18°C to a maximum of 34°C, with an average of 25.5°C, and the relative humidity ranges from 76% to 84%, with an average of 81% (Fig. 1). Capture sites and sand fly collection and identification - Sand fly collections were carried out in three patches of forest (patch A: 4°59'54.3"N, 72°42'39.6"W; patch B: 5°00'59.9"N, 72°42'54.3"W, and patch C: 5°00'46.8"N, 72°42'46.4"W) bordering small rivers. These alluvial gallery forests are a common element of the tropical grassland savannas (Llanos Orientales) in Colombia. The sand flies were collected during two periods of two consecutive years, with monthly collections of three nights from March 1996 to February 1997 and August 1997 to July 1998. To investigate the geographical distribution, activity, seasonality, and anthropophilic habits of the species, different methods of capture were used (Alexander 2000). The collections were performed by: (i) aspiration of sand flies from tree trunks and buttress roots to investigate diurnal resting places by one collector for about 5 min per tree, with a total of 250 and 222 trees included in the survey during the 1st and 2nd years, respectively; (ii) disturbing those sand flies resting on leaf litter during the day with a mesh-covered wooden frame containing a triangular cross-section with an area of 0.3 m2, an area of 9 m2/month was covered during the first year (this technique was not used in the 2nd year); (iii) using CDC miniature light traps suspended approximately 2 m above the ground each night, with two traps/night set 1 h before sunset and collections performed after sunrise the next morning; (iv) using one Shannon trap between 18:30 h and 20:30 h each night; (v) human landing collection of insects attracted to arms exposed by one collector during the Shannon trap sampling time (i.e., between 18:30 h and 20:30 h); and (vi) human landing collections during daylight hours, while aspiration of sand flies from tree trunks and leaf litter were carried out. All sand flies were preserved in 70% alcohol for transportation to Centro de Investigaciones en Microbiología y Parasitología Tropical, where they were cleared in lactophenol, prepared and identified to species (Young & Duncan 1994). Sand fly cryopreservation - Sand flies collected between August 1997 and January 1998 were cryopreserved in liquid nitrogen and later dissected in sterile PBS (pH 7.4) under a compound microscope according to standard methodology (Young et al. 1987). Midgut, hindgut, and Malpighian tubules were examined in order to detect flagellates. The presence, location, and intensity of flagellates were noted, as well as whether they were alive and active (Minter & Goedbloed 1971). Attempts to culture sandfly flagellates were made using NNN media (Minter & Goedbloed 1971, Young et al. 1987). Data analysis - The correlation between the number of sand flies captured and the year of sampling was analyzed using Spearman's rank correlation test (Zar 1999). SPSS software was used to perform linear regressions between the average monthly number of sand flies captured using all sampling methods during the two years and the following environmental variables: precipitation, maximum temperatures, and relative humidity. Percentage values of relative humidity were used after angular transformation (Zar 1999). RESULTS A total of 2,365 sand flies (845 males and 1,520 females) from two different groups, five subgenera, and 10 different species were collected (Table I). Lu. fairtigi represented the species with the highest number of individuals captured during both years (55%), followed by Lutzomyia micropyga (Mangabeira) (20.9%) and Lutzomyia antunesi (Coutinho) (13.5%). Lu. fairtigi was not only abundant in the alluvial gallery forests of CA, but was also found to be an anthropophilic species, with individuals captured during human landing collections at night, as well as cloudy days (around 9:00 h and 16:00-17:30 h). Lu. micropyga was captured exclusively at tree buttresses used as diurnal resting sites. Collections on leaf litter covered approximately 108 m2, and resulted in 15 sand flies of five species (Table I), with an average yearly density of one sand fly per 7 m2. Lu. fairtigi, Lu. antunesi, and Lutzomyia panamensis (Shannon) were the most frequently collected species when using CDC and Shannon light traps (Table I). A seasonal fluctuation in the density of Lu. fairtigi was found to be correlated with the average monthly rainfall, maximum temperature, and relative humidity (Table II). The same analysis for Lu. micropyga and Lu. antunesi did not demonstrate a correlation between sand fly densities and environmental variables (Table II). However, the mean monthly numbers of Lu. micropyga collected increased during the rainy months from April to October. The same trend was observed for Lu. fairtigi, which disappeared during the months without rainfall (January to March), while Lu. micropyga and Lu. antunesi were present throughout the year (Fig. 3). A multiple linear regression between the three most abundant species and the environmental variables (rainfall, maximum temperature, and relative humidity) showed a significant correlation for Lu. fairtigi (R2 = 0.796, F = 10.38 and p = 0.004). In the cases of Lu. antunesi and Lu. micropyga, no significant correlations with environmental variables were found (R2 = 0.104, F = 0.31, p = 0.818; R2 = 0.441, F = 2.106, p = 0.178, respectively). Sand flies representing eight Lutzomyia species were dissected: Lu. antunesi, Lu. damascenoi (Mangabeira), Lu. fairtigi, Lu. flaviscutellata (Mangabeira), Lu. micropyga, Lu. panamensis, Lu. punctigeniculata (Floch & Abonnenc), and Lu. walkeri (Newstead). A total of 328 females were examined for natural infection with flagellate forms, but only one out of 259 Lu. fairtigi females demonstrated abundant and very active free Leishmania-like forms in an area posterior to the pylorous, between the rectal sac and the anus (posterior gut). The flagellates were not present in the midgut or Malpighi tubules. An attempt to isolate flagellate forms in NNN medium was terminated 48 h after inoculation due to strong bacterial contamination. DISCUSSION Lu. fairtigi is an endemic species from Colombia that is distributed in four of its Eastern departments (Bejarano 2007). Nothing is known about the epidemiological importance of Lu. fairtigi, and non-natural infections with flagellate forms have been reported for this species (Young & Duncan 1994). Our study shows for the first time the presence of flagellate forms in one Lu. fairtigi female. If the flagellates detected in this single infected Lu. fairtigi were Leishmania parasites, the Leishmania natural infection rate for this species in the Tauramena Municipality would be of 0.3%. The peripyloric location (posterior gut) of the flagellates suggests that they belong either to the genus Leishmania subgenus Viannia or genus Endotrypanum (Christensen & Herrer 1976). Shaw (1992) distinguished infections caused by these two genera based on the abundance of flagellates in Malpighi tubules, in the case of Endotrypanum, and the absence of tubule infection, in the case of subgenus Viannia. However, Ramos et al. (1997) and Barbosa et al. (2006), discarded this criterion and, together with Christensen and Herrer (1976), highlighted individual variations in the intestinal distribution of Endotrypanum, with the greatest number of flagellates found in the midgut. Barbosa et al. (2006) found in experiments with Lutzomyia longipalpis that Endotrypanum and L. (V.) guyanensis differed significantly in relation to the colonization of the stomodeal valve. Williams (1999) describes both sexes of the Psychodopygus subgenus as canopy dwellers, with females also feeding in the canopy. Such behavior suggests the possibility of females being in contact with tree sloths, the vertebrate hosts of Endotrypanum (Shaw 1992). However, many Psychodopygus females have been found naturally infected with Leishmania species (Williams 1999, Young & Duncan 1994). Consequently, the presence of different flagellates in the mid and hindgut of sand flies (Johnson et al. 1963, McConnell 1963, McConnell & Correa 1964, Wallace & Hertig 1968) and unsuccessful attempt to isolate the flagellar forms found in one female do not answer questions associated with parasite identification and the potential vector status of Lu. fairtigi. Among the other captured species, the following are important for their vector status: (i) Lu. panamensis (vector of L. panamensis in Panama and a suspected vector in Venezuela and Honduras), and (ii) Lu. flaviscutellata (vector of L. amazonensis in Brazil, Colombia, and French Guyana, and a suspected vector in Ecuador, Surinam, Trinidad and Tobago, and Venezuela) (WHO 1990). The low anthropophilia of Lu. flaviscutellata and its strong preference for wild rodents (Oryzomys and Proechimys) and birds (Lainson & Shaw 1968) explains the rare transmission of Leishmania parasites to humans (Shaw & Lainson 1968, Rodríguez et al. 1985). Primary forests, depleted secondary forests, isolated vegetation spots, gallery forests, and "varzea" are some of the ecosystems where Lu. flaviscutellata has been reported (Shaw & Lainson 1972). However, L. amazonensis parasites were isolated from six human cases in the departments of ME and Northern Santander in Colombia (Corredor et al. 1990). The geographic distribution of these cases close to CA and the presence of Lu. flaviscutellata in Eastern Colombia (Bejarano 2007) suggest that the gallery forests in CA could be considered as an ecosystem involved in L. amazonensis transmission. Human cases caused by other Leishmania species have been reported in CA, and include L. panamensis, L. braziliensis (Saravia et al. 1998), L. mexicana, and L. mexicana complex (Ovalle et al. 2006). The presence of Lu. panamensis in this region may therefore be an important factor for the transmission of L. panamensis. The different collection methods (Table I) show how species like Lu. micropyga were exclusively captured on trees used as diurnal resting sites. This association has been related to the strong preference of Lu. micropyga to feed on cold-blooded vertebrates (WHO 1990). CDC traps captured a large number of Lu. fairtigi and Lu. antunesi individuals (Table I), demonstrating the characteristic canopy-floor displacement of females belonging to the Psychodopygus subgenus (Williams 1999). The presence of Lu. flaviscutellata on leaf litter highlights its relationship with low-growing shrubs as a diurnal resting site (Shaw et al. 1972), as well as a site that enhances the contact of females with their food source (rodents). Lu. fairtigi captured on leaf litter is in agreement with the observations of Chaniotis et al. (1972), who established that ground level capturing results in the collection of species with anthropophilic habits. Diurnal and nocturnal human landing collections, the high population density in gallery forests during months of rainfall, and the presence of flagellate forms between the rectal sac and the anus of Lu. fairtigi raise the question of the epidemiological importance of this endemic species to Eastern Colombia. ACKNOWLEDGEMENTS To Dr. Marta Wolff, for assistance with some literature, German Aguilera, for assistance with Fig. 1, and to referees for many detailed and helpful suggestions. REFERENCES
Copyright 2008 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc08084t1.jpg] [oc08084f2.jpg] [oc08084f3.jpg] [oc08084f1.jpg] [oc08084t2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}