 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
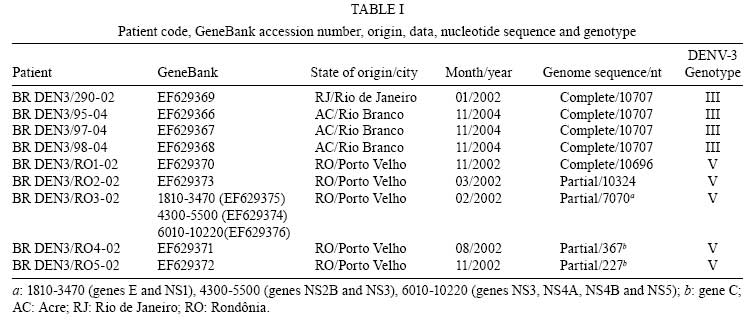
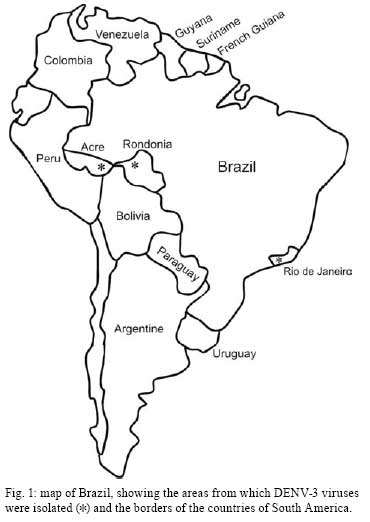
Memórias do Instituto Oswaldo Cruz, Vol. 103, No. 5, August 2008, pp. 483-488 Evidence for the co-circulation of dengue virus type 3 genotypes III and V in the Northern region of Brazil during the 2002-2004 epidemics Meri Bordignon NogueiraI, II, III; Vanessa StellaI; Juliano BordignonI; Weber Cheli BatistaIV; Luana de BorbaI; Luis Hildebrando Pereira da SilvaIV; Federico Guillermo HoffmannI; Christian Macagnan ProbstI; Claudia Nunes Duarte dos SantosI, * IInstituto
Carlos Chagas-Fiocruz, Rua Prof. Algacyr Munhoz Maeder 3.775, Cidade Industrial,
81350-010 Curitiba, PR, Brasil Financial support: Fiocruz, CNPq (proc. 410593/2006-0), Fundação Araucária (proc. 5-1-8892), CYTED/RIVE network Received 11 March
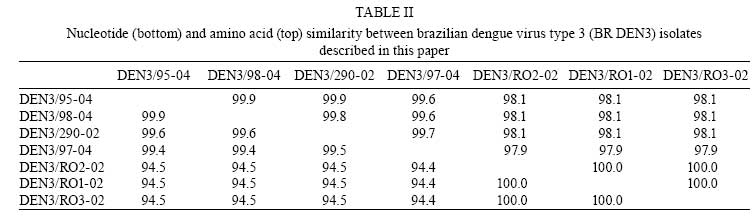
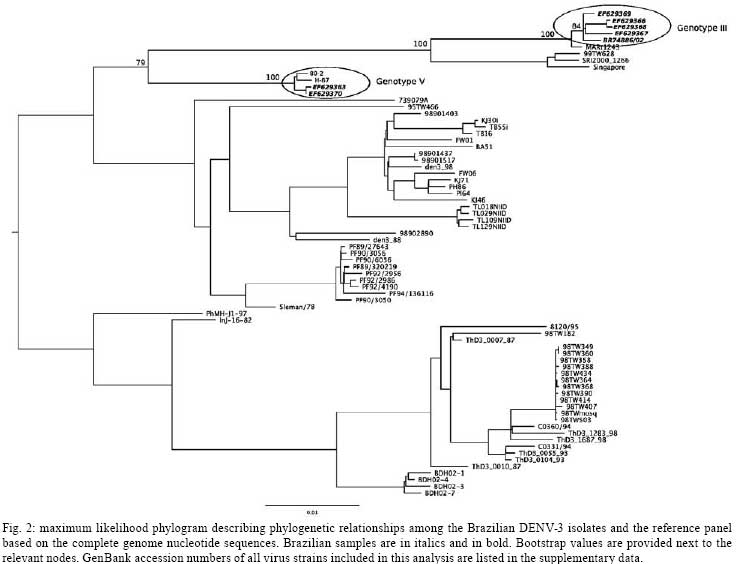
2008 Code Number: oc08085 ABSTRACT The reintroduction of dengue virus type 3 (DENV-3) in Brazil in 2000 and its subsequent spread throughout the country was associated with genotype III viruses, the only DENV-3 genotype isolated in Brazil prior to 2002. We report here the co-circulation of two different DENV-3 genotypes in patients living in the Northern region of Brazil during the 2002-2004 epidemics. Complete genomic sequences of viral RNA were determined from these epidemics, and viruses belonging to genotypes V (Southeast Asia/South Pacific) and III were identified. This recent co-circulation of different DENV-3 genotypes in South America may have implications for pathological and epidemiological dynamics. Key words: dengue virus type 3 - genotype III and V - co-circulation - phylogenetic analysis - Amazon Dengue is an emerging infectious disease affecting almost 50 million people in tropical and subtropical regions of the world. Dengue fever (DF) and dengue hemorrhagic fever (DHF) are caused by four closely related, but antigenically different dengue virus serotypes 1-4 (DENV-1-4) (Gubler & Clark 1995). Since the introduction of the DENV into Brazil in 1986, more than four million cases of dengue have been reported (Cordeiro et al. 2007), and the incidence of severe clinical symptoms has increased. No vaccine or specific therapeutic antiviral measures are available. Several studies have suggested that individuals experiencing a second dengue infection with a heterologous serotype are at greater risk for developing DHF or dengue shock syndrome (DSS) (Cummings et al. 2005, Halstead et al. 2005). However, DHF and DSS are also observed in primary cases, and not all secondary infections correspond to severe forms of the disease. Studies based on molecular epidemiology have provided evidence that differences in virulence between viral strains could play a role in the severity of the disease (Mangada & Igarashi 1998, Rico-Hesse 2003). Major dengue virus epidemics occurred in Brazil in 1986 and 1990, due to the introduction of DENV-1 and -2, respectively, and the subsequent spread of these serotypes throughout the country. DENV-3 reappeared in 1994 in Central America (Nicaragua and Panama) after 17 years of absence, and later spread to Mexico and the Caribbean (CDC 1995, Guzman et al. 1996, Usuku et al. 2001). In 2000, the presence of this serotype was reported in two countries in South America: Brazil and Venezuela (Nogueira et al. 2001, Uzcategui et al. 2003). DENV-3 was initially isolated in December 2000 in the state of Rio de Janeiro (RJ), in Southeastern Brazil (Nogueira et al. 2005). The virus has since been detected in almost all Brazilian states, establishing conditions of hyperendemicity with serotypes 1, 2 and 3. DENV-3 genotype III (Sri Lanka/India) was identified in all affected states during the epidemics of 2001 to 2004, and caused the one of most severe epidemic with the largest number of reported cases, greatest severity of clinical manifestations in primary infections in particular, and the largest number of confirmed deaths (Nogueira et al. 2005). In 2002, Brazil contained almost 80% of the one million cases of dengue infection in the Americas, with almost 800,000 dengue cases and 150 deaths attributed to DENV-3 (www.saude.gov.br). Recently, Figueiredo et al. (2008) reported the circulation of DENV-3 genotype I in Minas Gerais. In the Amazonian state of Rondônia (RO), located in the Northern region of Brazil, the first confirmed cases of dengue dated from 1997, and the first outbreak occurred in 2000, with 2,759 cases (http://portal.saude.gov.br/portal/arquivos/pdf/taxa_incidencia_dengue2007.pdf). DENV-1 was isolated in 2001 in Porto Velho (the largest city in the state). In 2002, following the introduction of DENV-3, the state experienced an epidemic, in which unusual clinical symptoms, such as meningoencephalitis, were observed (Nogueira et al. 2005, WC Batista, unpublished observations). Given the high incidence of severe clinical symptoms related to primary DENV infections observed in the 2002 epidemic, we decided to characterize the genome of the virus circulating in the Northern region of Brazil and determine its phylogenetic relationship with other DENV-3 strains. Identification of the circulating DENV genotypes is important, as it has been demonstrated that some strains are more frequently associated with severe disease than others (Leitmeyer et al. 1999, Pandey & Igarashi 2000, Rico-Hesse 2003). PATIENTS, MATERIALS AND METHODS Clinical samples and viruses - A serum sample from a dengue positive patient who had traveled to an area of epidemic dengue in RJ in Southeastern Brazil, and eight serum samples from patients living in Acre (AC) and RO in Northern Brazil (Table I, Fig. 1) were kindly provided by LACEN, Curitiba and IPEPATRO, Porto Velho, respectively. All patients were diagnosed with DF. These samples were used for the characterization of DENV genomes. RNA extraction and RT-PCR - Viral RNA was isolated from 140 µ of patient serum samples, taken during the acute phase of the disease and/or from 140 µ of purified virus prepared from cell culture supernatants after three passages in C6/36 cells, as described elsewhere (Duarte dos Santos et al. 2002). The QIAamp Viral RNA Mini Kit (Qiagen, Valencia, USA) was used, following the manufacturer's instructions. Complete genomes were amplified by means of overlapping RT-PCR products, using 5 l of RNA and random primers (Invitrogen, Carlsbad, USA) and Improm II Reverse Transcriptase (Promega, Madison, USA), for 1 h at 47 ºC; 5 µ of cDNA were used for the amplification reaction with the High Fidelity TripleMaster® PCR System (Eppendorf, Hamburg, Germany) and specific primers, according to the protocols supplied with the kit. Briefly, the thermocycling conditions consisted of 94ºC/3 min, followed by 35 cycles of 94ºC/30 s, 55-58ºC/30 s and 68ºC/3 min. Sequencing strategy, multiple sequence alignment and phylogenetic analysis - The amplicons were directly sequenced using a Thermo Sequenase kit (USB Inc, Ohio, USA) on an ABI3100 device, with the BigDye7 Terminator method (Applied Biosystems, Warrington, UK). Nucleotide sequences were analyzed with a Phred/Phrap/Consed package (www.phrap.org). Sequences were obtained from the 5' and 3' UTR of the BR DEN3/290-02, BR DEN3/95-04, BR DEN3/97-04, BR DEN3/98-04, BR DEN3/RO1-02 and BR DEN3/RO2-02 samples after uncapping and RNA ligation, as described elsewhere (Duarte dos Santos et al. 2000). Primer sequences are available upon request. Sequence analyses - For phylogenetic and comparative purposes, we assembled a representative panel of complete DENV-3 genome sequences from GenBank (supplementary data, Supp 1, 2). All sequences were aligned using CLUSTALW (Thompson et al. 1994). Phylogenies were estimated in a Maximum likelihood framework using Treefinder (version April, Jobb et al. 2004). We selected the best-fit model of nucleotide substitution based on the Bayesian Information Criterion using the Model Proposal routine from Treefinder. Support for the nodes was evaluated by running 500 bootstrap pseudoreplicates. The likelihood scores of competing hypotheses were obtained using the Shimodaira-Hasegawa (SH) topology test (Shimodaira & Hasegawa 1999), as implemented in Treefinder (Jobb et al. 2004). RESULTS In order to characterize the DENV strains circulating in Brazil and determine their relationships with other DENV strains, we examined clinical samples from RO (epidemic in 2002) and AC (epidemic in 2004), and from RJ (epidemic in 2002), located in the Northern and Southeastern regions of Brazil, respectively. Nine DENV isolated from human samples collected in these three different geographical areas of Brazil were sequenced after three passages in C6/36 cells. Complete genome sequences were determined for five of these viral isolates: BR DEN3/290-02, BR DEN3/95-04, BR DEN3/97-04, BR DEN3/98-04 (alignment of 10707 nt) and BR DEN3/RO1-02 (alignment of 10696 nt). In addition, an almost complete genome sequence of the BR DEN3/RO2-02 virus (starting at position 378 and ending at position 10696), as well as three fragments covering ~70% of the genome from the BR DEN3/RO3-02 strain (corresponding to positions 1810-3470, 4300-5500 and 6010-10220) were determined. The BR DEN3/RO1-02 and BR DEN3/RO2-02 sequences had an 11 bp deletion in the 3´UTR. We also determined partial genomic sequences from viral RNA extracted directly from the plasma of two Rondônian patients (BR DEN3/RO5-02, from nt 135 to 361 and BR DEN3/RO4-02, from nt 132 to 498 of DENV-3 RNA). Both samples were identified as belonging to genotype V. We compared complete genomic sequences between five dengue viruses and 7070 nt between seven dengue viruses (due mainly to the fragmentation of BR DEN3/RO3-02). Their nucleotide and amino acid similarities are displayed in Table II. Comparisons of complete genome nucleotide sequences showed that BR DEN3/290-02 was most similar to BR74886/02 (99.6% identity, AY679147), which was isolated from the liver of a patient who died from the disease in RJ (Miagostovich et al. 2002, 2006). BR DEN3/RO1-02 was found to be closely related to 80-2 (99.7% identity, AF317645), a virus from a DF patient isolated in 1980 in China. As can be seen in Fig. 2, the samples from RO formed a separate cluster, which was closely related to the DENV-3 H-87 (M93130), D3-73NIID (AB111085) and 80-2 (AF317645) isolates, which belong to genotype V (Southeast Asia/South Pacific), and are clearly different from those from AC and RJ isolates, which clustered with genotype III (Sri Lanka/India, AY099336). This finding is remarkable, as the three genotype V viruses were isolated in Asia in 1956, 1973 and 1980, respectively. Brazilian DENV-3 isolates grouped into two separate clides: all samples isolated from Acre grouped with an isolate from RJ, whereas all samples isolated from RO were clustered in a separate clade (Fig. 2). Bootstrap support for nodes separating the two groups of Brazilian samples was strong (Fig. 2), and a tree where all sequences from Brazil were clustered in a single group providing significantly worse fit to the data (SH test p < 0.001). DISCUSSION A displacement of serotypes 1 and 2 of DENV was documented in Brazil after the introduction of a highly virulent genotype of DENV-3 (genotype III, Sri Lanka/India) in 2002 (Nogueira et al. 2005). The rapid spread of DENV-3 caused severe epidemics in almost all Brazilian regions in the three years following its introduction (Nogueira et al. 2005, De Simone et al. 2004). The Northern region of Brazil suffered DENV-3 epidemics between 2002 and 2004. Different clinical profiles were observed during this epidemic period, with DHF/DSS and encephalitis cases identified in RO, whereas only DF was observed in AC. During the DENV-3 epidemic in RO in 2002, several atypical encephalitis cases were observed in patients testing positive serologically for dengue, but it was not possible to isolate virus from those patients. It remains unclear whether that clinical pattern was exclusively related to DENV-3, or was the result of co-infection with other viruses, such as the SLE or Rocio viruses (Rocco et al. 2005, Mondini et al. 2007). However, we cannot exclude the possibility that the observed unusual clinical presentations were exclusively due to the DENV infections, taking into account the results of Domingues et al. (2008) which showed that 21.2% of patients with dengue infection exhibited involvement of the central nervous system. In AC, DENV-3 was first isolated in 2004 and no severe case was observed at that time (Brazilian Ministry of Health, SVS, www.saude.gov.br). We compared viral genomes amplified directly from serum samples or from insect cells infected with viruses circulating in Northern Brazil, samples of DENV-3 from RJ, and those retrieved from databases. It should be emphasized that only DENV-3 genotype III was identified during the 2002 to 2003 epidemic in samples from Porto Velho (Aquino et al. 2006) and RJ (Nogueira et al. 2005), and this genotype is the only one to have been implicated in DENV-3 epidemics in the Americas (Usuku et al. 2001, Messer et al. 2003, Uzcategui et al. 2003, Peyrefitte et al. 2005, Rigau-Pérez & Laufer 2006, Ocazionez et al. 2006). When samples received from Porto Velho were used to infect C6/36 cells, they displayed a cytotoxic effect not seen with other DENV-3 viral isolates (data not shown). These findings led to the molecular characterization of these samples. Viral genome sequences of five isolates indicated that DENV-3 genotypes III and V co-circulated during the 2002 epidemic in RO. We analyzed three samples from AC, all of which were of genotype III, as well as the isolate from RJ. However, due to the small number of samples studied, we cannot rule out the possibility that another genotype was also circulating in those regions. The virus collection of our institute (ICC-Fiocruz) does not include DENV-3 strains H87 (M93130, Osatomi & Sumiyoshi 1990), 80-2 (AF317645), or any other DENV-3 genotype V strain, and these strains had never been manipulated in our laboratories. Sample contamination was therefore highly unlikely. Furthermore, two of the eight samples from RO and seven of the 12 samples from AC tested negative. Moreover, viral RNA for BR DEN3/RO4-02 and BR DEN3/RO5-02 was obtained directly from plasma samples. It is worth mentioning that our data was recently corroborated by similar findings in Colombia (José Usme Ciro and Juan Carlos Gallego-Gómez, unpublished observations) which showed that DENV-3 genotype V had also been isolated from human cases in Colombia during the 2003-2005 epidemics. Aquino et al. (2006) recently demonstrated by phylogenetic analysis with partial nucleotide sequences for the E protein and 3'UTR that Brazilian DENV-3 (particularly from RO) are grouped with samples from Sri Lanka, Samoa and other American genotype III DENV-3. These authors also suggested that DENV-3 had been introduced at least twice into Brazil, once via RJ and the second via the Caribbean countries. More recently, Figueiredo et al. (2008) demonstrated the co-circulation of two genotypes of DENV-3 in Minas Gerais from 2002 to 2004. A larger number of viral isolates should be analyzed in other Brazilian states and South American countries to identify the circulating dengue genotypes, and to track the dynamics of virus introduction and maintenance in nature. The DENV-3 genotype classification is a controversial matter. Wittke et al. (2002) have re-classified the China 80-2 strain, formerly classified as genotype I, as genotype V based on E gene sequences. Aquino et al. (2006) have followed this classification. On the other hand, Figueiredo et al. (2008), using C/prM gene sequences, classified the same strain as belonging to genotype I. We decided to adopt the Wittke et al. (2002) classification as it provided a higher level of genotype discrimination. Following this, we classified our strains as belonging to genotypes III (AC) and V (RO). However, we acknowledge that DENV-3 genotype classification is still controversial. Intra-serotype genetic diversity in DENV vary greatly on a temporal scale, and it has became clear that individual lineages or entire virus clades frequently arise, persist for a period of time, and then disappear (Zhang et al. 2005). Therefore, more precise criteria for genotype classification of dengue virus are essential to avoid different nomenclature for similar strains. The results obtained in this study raise some intriguing questions: how can a recent clinical viral isolate from South America display 99.9% identity with a laboratory viral strain (DENV-3 80-2) isolated from a DF patient in China? Will it be possible to trace the route by which these viruses entered the Americas? Interestingly, the DENV-3 genotype V strains H-87 (L11423) isolated from a human case in the Philippines in 1956, D3-73NIID (AB111085) isolated in 1973 from an imported human case in Japan, and 80-2 (AF317645) isolated in China in 1980, also display a remarkably high level of nucleotide sequence conservation (over 99%) with each other and with some of the viruses included in this study. The most dramatic case of sequence similarity is illustrated by the three viral isolates from RO; they display 100% sequence identity with each other. These three cases are probably linked epidemiologically, but further efforts are needed to elucidate whether they represent a single chain of infection. Further genome characterization of viruses from other South American countries that have also experienced unusual DENV-3 epidemics should shed light on this issue. ACKNOWLEDGEMENTS To Paulo Arauco for technical help with sequencing and Anaclete Felinni, from LACEN-Curitiba, for the dengue serum samples. REFERENCES
Copyright 2008 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc08085f1.jpg] [oc08085t2.jpg] [oc08085t1.jpg] [oc08085s1.jpg] [oc08085s2.jpg] [oc08085f2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}