 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
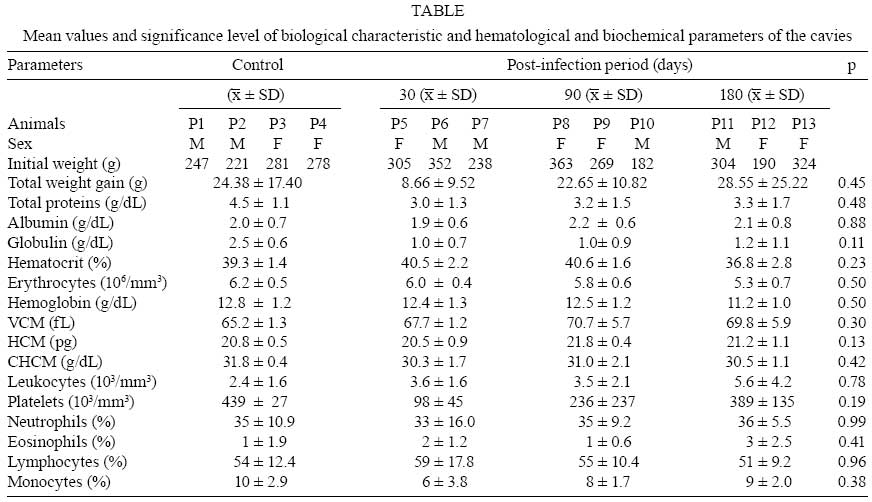
Memórias do Instituto Oswaldo Cruz, Vol. 103, No. 6, September, 2008, pp. 545-548 Experimental infection parameters in Galea spixii (Rodentia: Caviidae) with Leishmania infantum chagasi Patrícia Batista Barra Medeiros BarbosaI; Paula Viviane Sousa de QueirozI; Selma Maria B JerônimoII; Maria de Fátima Freire de Melo XimenesIII, + IFaculdade
de Ciências da Saúde, Universidade Estadual do Rio Grande do Norte,
Natal, RN, Brasil Financial support: CNPq, CAPES Received 24 March
2008 Code Number: oc08099 ABSTRACT In order to better understand the epidemiological transmission network of leishmaniasis, an endemic disease in Northeast Brazil, we investigated the susceptibility of Spix yellow-toothed cavies (Galea spixii) to the Leishmania infantum chagasi parasite. Nine cavies were experimentally infected, separated into three groups and monitored at 30, 90 and 180 days, respectively. Amastigotes were identified in the spleen slides of two cavies killed 180 days after infection. Antibodies against the L. i. chagasi were identified in one of the cavies. This demonstrates that G. spixii is in fact capable of maintaining a stable infection by L. i. chagasi without alterations in biochemical and hematological parameters of the host and without perceivable micro and macroscopic lesions. Key words: Galea spixii - Leishmania infantum chagasi - animal hosts American visceral leishmaniasis (AVL) remains an important public health problem in Brazil. The existence of wild reservoirs, many still unknown, has compromised the control measures for this zoonosis, which currently consist of only the elimination of parasite-positive dogs, the main urban reservoirs of Leishmania infantum chagasi (Lainson et al. 2002). Several species of rodents are suspected to be the principle wild reservoirs of the Leishmania species responsible for the tegumental form of the disease. Cases of natural infection by L. i. chagasi registered in Rattus rattus (Zuleta et al. 1999, Oliveira et al. 2005), in Trychomys apereoides (Oliveira et al. 2005) and Proechimys canicollis (Travi et al. 1998) illustrate the need for a better understanding of the role of these animal reservoirs in AVL transmission. Spix yellow-toothed cavies (Galea spixii Wagler) are small rodents belonging to the suborder Hystricognathi, family Caviidae, subfamily Caviinae. They are tailless and have a dark gray dorsal surface and white ventral surface with infraocular and postauricular white spots. These rodents are morphologically similar to the other Caviinae species; however, their differentiating characteristic is their yellow incisors (Lacher 1981). They are found in all the states of Northeast Brazil. Although studies have been carried out to address raising these animals in captivity (Pinheiro et al. 1989), little research has been done on the zoonoses that they might transmit. The purpose of this study was to investigate the susceptibility of Spix yellow-toothed cavies to the L. i. chagasi parasite through the experimental infection of these animals in order to better understand visceral leishmaniasis epidemiological chains in Northeast Brazil. MATERIALS AND METHODS The experimental protocol was approved by the Research Ethical Committee of the Universidade Federal do Rio Grande do Norte. Permission to capture 21 Spix yellow-toothed cavies (Figure) was given by the Instituto Brasileiro do Meio Ambiente e dos Recursos Naturais Renováveis. The animals were captured in peripheral areas of Mossoró, a city located in the western part of the state of Rio Grande do Norte, Northeast Brazil. After clinical examination, the cavies were sorted randomly and housed in metal cages (80 x 45 x 60 cm) which hold a maximum of four animals each. The cages contained feeders, water dispensers and shelters and were kept isolated in a controlled room. Food was provided ad libitum and replaced daily. It consisted of rodent ration (Labina, Purina) complemented with sweet potato, leafy greens [lettuce, carrot leaves and chopped elephant grass (Pennisetum purpureum)] and green beans. After a 40-day period of observation and adaptation to the experimental conditions, 13 cavies were allocated to four groups, with G1 (4 animals) as the control. The animals of the remaining groups (G2, G3 and G4) were intraperitonially inoculated with 107 promastigotes of L. i. chagasi obtained from the spleen and liver of naturally infected dogs and monitored for 30, 90 and 180 days, respectively. To control parasite virulence, two hamsters were infected under the same conditions as the cavies. All animals in all groups were clinically assessed once a week. The parameters adopted included the following: skin elasticity, the color of the buccal and conjunctive mucosas, abdominal sensitivity and body coat. Rectal temperature and total weight gain were also measured. After the post-infection period, the animals were anesthetized with an intramuscular injection of 75 mg/Kg of tiletamine chloride and zolazepam chloride (Zoletil 50). Blood collected by cardiac punction was used to perform a complete hemogram with an automatic cell counter (Cell-dyn 1400) and a differential leukocyte count in blood smears stained by the Panotico method (methylene blue, eosin, ethyl alcohol, Laborclin) and to obtain serum to determine total protein and albumin values using commercial tests (Labtest) and anti-Leishmania antibody studies. Peripheral blood aliquots, as well as fragments of the liver, spleen, popliteal and mesenteric lymph nodes, were used to prepare Giemsa-stained slides and biphasic culture medium (NNN/Schneider). The cultures were incubated at 24ºC for four weeks and observed daily. Formaldehyde spleen fragments were used to prepare histological slides stained with Giemsa and hematoxylin-eosin. Given the small spleen size, after preparation of slides, culture and histopathological examination, there was no material left for DNA extraction and subsequent parasitical study. For this reason, the procedure (DNA extraction) was performed only on the liver. A 100 mg fragment of liver homogenized in 1.2 mL of TE solution (10 mM Tris-CI, pH 8.0; 0.1 mM EDTA, pH 8.0) was digested at 55ºC for 2 h in 40 µL of 20% SDS and 60 µL of proteinase K (20 mg/mL). After successive extractions with phenol, isoamylic phenol/chloroform/alcohol (25:24:1) and isoamylic chloroform/alcohol (24:1) and precipitation with two volumes of ethanol, the DNA was resuspended in 200 µL of TE and incubated at 37ºC for 30 min before being frozen at -20ºC. The polymerase chain reaction (PCR) was carried out using complementary oligonucleotides of kinetoplast minicircle sequences from Leishmania as described by Smyth et al. (1992). To the reaction we added 20 ng of DNA extracted at a mixture of 4.185 µL of bidistilled sterile water, 1.25 µL of PCR buffer (100 mM Tris-HCI, 500 mM KCI, 15 mM MgCl2, pH 8.3), 1.25 µL of each oligonucleotide, at a concentration of 20 nmol (right -5' GGG GTT GTA AAA TAG and reverse -5' CCA GTT TCC CGC CCC G), 2.5 µL of a solution containing 1.25 mM desoxyribonucleoside triphosphate and 0.0625 µL of 5 U/µL of thermo-resistant DNA polymerase. The samples were initially denatured at 93ºC for 30 s. Annealing occurred at 60ºC for 1 min, and the final extension at 72ºC for 1 min. Identification of the PCR product was performed by agarose gel electrophoresis The study of anti-Leishmania antibodies in the infected animals was performed by means of immunoenzyme trial, using the recombinant rK39 protein of L. i. chagasi (Qu et al. 1994) and the total extract of L. i. chagasi (raw extract). Peroxidase-stained protein-A (protein HRP conjugate grade/CN biomed OH44202) was diluted in 0.1% PBS-Tween 20 at 1:10000 (for the rK39 antigen) and 1:1000 (for the raw extract). Serum from AVL-positive dogs was used as control and to obtain cutoff points (0.159 for rK39 and 0.050 for the raw extract); we used serum from the control group animals (G1) and from four cavies born in the laboratory during the study. To compare the means of the parameters analyzed, we used the Kruskal-Wallis non-parametric test and Statistica software, version 6.0, at a significance level of 5%. RESULTS At the moment of capture, no clinical sign suggestive of leishmaniasis or of any other infirmity was observed in the cavies. During the 40-day observation period, seven animals died and necropsy observations showed no lesions compatible with leishmaniasis. Another cavy was eliminated from the experiment for having undergone antibiotic treatment for wounds received in a fight. During monitoring after L. i. chagasi infection, a wide variation in total weight gain was observed among the animals. However, there were no significant differences between the means of the different groups. Rectal temperature also did not differ statistically between control and infected animals. Considering the controls and the infected animals, the amplitude and the temperatures of all animals ranged from 36-40ºC. The mean rectal temperatures (mean ± standard deviation) observed in the control group animals and in those infected for 30, 90 and 180 days were 38.1ºC ± 0.7; 37.3ºC ± 0.5; 37.9ºC ± 0.5 and 38.3ºC ± 0.6, respectively. No differences were found in the biochemical and hematological parameters of the groups (Table). Sparse amastigote forms of Leishmania were visualized in apposition on the slides and in the histological examination of the spleens of two females, both belonging to the group monitored for 180 days, and anti-Leishmania antibodies were also found in one of them. However, no pathological alterations were observed in the spleen of the animal, upon either macroscopic observation or histological examination. The DNA of L. i. chagasi was not found in all of the liver samples from all of the animals submitted to PCR. DISCUSSION The present study investigated the susceptibility of Spix yellow-toothed cavies (G. spixii) to the L. i. chagasi parasite by means of experimental infection. These small rodents, by virtue of their good adaptation to the inhospitable conditions of the semiarid region of Northeast Brazil, are generally found near residences and on the outskirts of rural cities (Lacher 1981, Pinheiro et al. 1989). No differences in total weight gain and rectal temperature were observed between the animals infected and the controls. Complete hematocrit values and the biochemical parameters of total proteins (albumin and globulin) did not differ between the groups either, most likely because of the absence of severe infection causing systemic damage in the parasite-inoculated animals. It should be pointed out that this is the first report of hematological and biochemical parameters in Spix yellow-toothed cavies. The presence of scarce parasites and the absence of significant pathological alterations indicate a sub-patent infection with low parasite load. Although there is currently no concrete evidence of natural L. i. chagasi infection in cavies, cutaneous lesions attributed to leishmanias have been recorded in another cavy species (Cavia porcellus), without identification of the species of Leishmania (Ecco et al. 2000) or have been identified and attributed to Leishmania enrietti (Medina 1946, Muniz & Medina 1948, Machado et al. 1994). To understand the infection in only two animals in this study, even though all animals were submitted to the same experimental condition, one must consider the genetic heterogeneity of the wild population, which implies the existence of different levels of susceptibility among the individuals, a fact that has been well established in murine models (Blackwell 1996). This difficulty in experimentally infecting cavies suggests, even without knowledge of the cytokine profile produced, that cavies are resistant to L. i. chagasi infection. However, G. spixii are abundant, highly prolific animals, living in proximity to humans and dogs and capable of maintaining a stable infection without alterations in biochemical and hematological parameters and even without perceivable macro and microscopic lesions. Thus, we suggest further studies to help clarify the role of G. spixii cavies as hosts for L. i. chagasi in Northeast Brazil. REFERENCES
Copyright 2008 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc08099t1.jpg] [oc08099p1.jpg] |
| |||||||||

{kind=link}
{kind=link}