
|
Memórias do Instituto Oswaldo Cruz
Fundação Oswaldo Cruz, Fiocruz
ISSN: 1678-8060 EISSN: 1678-8060
Vol. 103, Num. 7, 2008, pp. 629-644
|
Memórias do Instituto Oswaldo Cruz, Vol. 103, No. 7, November, 2008, pp. 629-644
A
REVIEW
The utility
of rhesus monkey (Macaca mulatta) and other non-human primate models
for preclinical testing of Leishmania candidate vaccines
Gabriel Grimaldi
Jr
Laboratório
de Pesquisas em Leishmaniose, Instituto Oswaldo Cruz-Fiocruz, Av. Brasil 4365,
21045-900 Rio de Janeiro, RJ, Brasil
Corresponding author: grimaldi@ioc.fiocruz.br
Financial support: Millennium Institute for Vaccine Development and Technology
(MCT/CNPq-420067/2005-1).
Received 15 October
2008
Accepted 22 October 2008
Code Number: oc08116
ABSTRACT
Leishmaniasis causes
significant morbidity and mortality, constituting an important global health
problem for which there are few effective drugs. Given the urgent need to identify
a safe and effective Leishmania vaccine to help prevent the two million
new cases of human leishmaniasis worldwide each year, all reasonable efforts
to achieve this goal should be made. This includes the use of animal models
that are as close to leishmanial infection in humans as is practical and feasible.
Old world monkey species (macaques, baboons, mandrills etc.) have the closest
evolutionary relatedness to humans among the approachable animal models. The
Asian rhesus macaques (Macaca mulatta) are quite susceptible to leishmanial
infection, develop a human-like disease, exhibit antibodies to Leishmania
and parasite-specific T-cell mediated immune responses both in vivo and in vitro,
and can be protected effectively by vaccination. Results from macaque vaccine
studies could also prove useful in guiding the design of human vaccine trials.
This review summarizes our current knowledge on this topic and proposes potential
approaches that may result in the more effective use of the macaque model to
maximize its potential to help the development of an effective vaccine for human
leishmaniasis.
Key words:
non-human primates - experimental leishmaniasis - Leishmania vaccine
development
Leishmaniasis is
one of the major infectious diseases primarily affecting some of the poorest
regions of the world. The disease is endemic in 88 countries, and the World
Health Organization estimates that it is a threat to 350 million people with
a worldwide prevalence of 12 million cases. Among the annual incidence of 2
million new cases of human infections, 0.5 million are life-threatening visceral
leishmaniasis (VL) (www.who.int/tdr/diseases).
Cutaneous leishmaniasis (CL) caused by highly pathogenic parasites is also characterized
by its chronicity, latency and tendency to metastasize, resulting in recurrent
skin lesions with the potential for mucosal involvement. It should be noted
that an estimated 2.4 million disability adjust life years, in addition to 59,000
lives, were lost to leishmaniasis in 2001 alone (Davies et al. 2003). Concerns
about chemotherapy failure for both VL and CL are exacerbated by geographical
variation in antimonial treatment regimens, severity of disease and sensitivity
of Leishmania species. In addition, no proven successful vaccine for
controlling human leishmaniasis is in routine use (Davies et al. 2003, Kedzierski
et al. 2006). Moreover, at least 20 genetically heterogeneous Leishmania
species infect humans and each of them has a unique epidemiological pattern,
such that two or more parasite species are often sympatric in sylvan areas of
the Neotropics (Grimaldi & Tesh 1993). These data explain the limited success
of current control strategies based on conventional measures (such as vector
reduction and elimination of infected reservoir) for American leishmaniasis.
The solid protective
immunity observed in humans following convalescence to CL formed the basis for
practice of active immunization, beginning with deliberate inoculation of virulent
organisms ("leishmanization") in centuries past and continuing with vaccination
using a crude antigen preparation obtained from inactivated ("killed") promastigotes
of one or various species of Leishmania, formulated either with or without
BCG (bacillus of Calmette and Guerin) as an adjuvant (Grimaldi 1995). While
accumulated experience with mass vaccination both in the ex-USSR and in Israel
has clearly shown that a virulent strain of Leishmania must be used for
vaccination to succeed (Gunders 1987), several Phase III trials testing the
potential efficacy of various crude vaccine approaches have given conflicting
results. Overall, the results vary from 0-75% efficacy against CL and little
(< 6%) or no protection against VL (Grimaldi 1995, Coler & Reed 2005).
Although host genetics can have dramatic effects on T-cell responses to existing
vaccines (Lambert et al. 2005), several technical problems (including inadequate
information about the quality, stability and potency of the antigens) may provide
explanation for some of the variation in efficacy observed in those human vaccine
studies. Nevertheless, most experts believe that a preventive vaccine will be
essential if this disease is ever to be controlled worldwide (Coler & Reed
2005, Tabbara 2006, Kedzierski et al. 2006, Palatnik-de-Souza 2008, Silvestre
et al. 2008).
The current effort
to develop improved vaccines for leishmaniasis has led to the need for appropriate
animal models in which to test candidate vaccines (Hein & Griebel 2003).
There are reminders that the results from rodent models do not automatically
translate to humans (MacGregor et al. 1998). The use of non-human primates (NHP)
as animal models for the study of human diseases (including immunological studies
and drug and vaccine-development studies against infectious diseases) has become
increasingly important (Campos-Neto et al. 2001, Delgado et al. 2005, Giavedoni
2005, Gibbs et al. 2007, Nikolich-Žugich 2007, Souza-Lemos et al. 2008).
For instance, the SIV-macaque model is widely used for testing vaccine and therapeutic
strategies prior to conducting human clinical trials (Nathansson et al. 1999,
Hu 2005). This review aims to provide insight into the current knowledge on
vaccine studies against leishmaniasis, with emphasis on studies involving vaccination
and experimental infection in monkeys.
Vaccine studies
against leishmaniasis
A major international
research effort over the past 20 years has resulted in the identification of
various Leishmania antigen candidates for second and third-generation
vaccines (Coler & Reed 2005, Palatnik-de-Souza 2008). Information about
a multitude of immunization approaches representing all of the major vaccine
design strategies, including vaccines using live genetically attenuated parasites,
subunit proteins/peptides in adjuvants, naked DNA and infectious vectored vaccines
expressing genes coding for specific leishmanial antigens and combinations thereof
has been given in recent review articles (Coler & Reed 2005, Tabbara 2006,
Kedzierski et al. 2006, Palatnik-de-Souza 2008, Silvestre et al. 2008). Many
of these vaccines have been tested for immunogenicity and protective efficacy
in a variety of experimental models (such as inbred laboratory rodents, dogs
and NHP). Depending on the particular vaccine approach and animal model used,
varying degrees of protective immunity have been achieved, as determined by
the level of parasite burden in infected sites and/or lesion size following
infectious challenge.
Vaccination strategies
are based on the immunology of Leishmania infection (Vanloubbeeck &
Jones 2004, Von Stebut 2007). On the basis of compelling evidence that both
CD4+ (including multifunctional Th1 cells and central memory CD4+
T-cells) and CD8+ T-cells are key players in the immune response
to leishmaniasis (Reed & Scott 2000, Zaph et al. 2004, Darrah et al. 2007),
the scientific community has focused considerable efforts on the development
of prophylactic vaccines that elicit T-cell responses (Rhee et al. 2002, Tapia
et al. 2003, Sharma et al. 2006, Dondji et al. 2008) with the premise that such
interventions will confer protective effects in these conditions. In this regard,
sustained protective immunity against both murine CL and VL has been achieved
by DNA vaccines encoding antigen candidates (Gurunathan et al. 2000, Mendez
et al. 2001, Campos-Neto et al. 2002, Zanin et al. 2007, Dondji et al. 2008)
or leishmanial recombinant protein(s) formulated with improved vaccine adjuvants
(Pashine et al. 2005), including cytosine phosphate guanosine oligodeoxynucleotides,
CpG ODN (Rhee et al. 2002, Iborra et al. 2005) and cationic distearoyl phosphatidylcholine
(DSPC) liposomes (Bhowmick et al. 2007). Of note, long-term immunity elicited
by those vaccines corresponded to, in addition to the presence of leishmania-specific
Th1, CD8+ T-cells responses (Gurunathan et al. 2000, Rhee et al.
2002, Sharma et al. 2006). Additionally, heterologous prime-boost vaccination
regimes, such as combining DNA priming with a live vectored boost (Gonzalo et
al. 2002, Ramiro et al. 2003), or two different live vectors to prime and boost
a response (Dondji et al. 2005, Ramos et al. 2008) have been explored as a means
of raising protective T-cell responses (Hu 2005).
Due to the genetic
variability of human T-cell responses (across HLA haplotypes), T-cell vaccines
can elicit variable protective immunity (Robinson & Amara 2005). A second
limitation of T-cell vaccines is the potential for T-cells to become exhausted
by high levels of persisting antigens (Kostense et al. 2002). Another challenge
is the ability of leishmanial parasites to modulate their antigens to evade
immune responses (Vanloubbeeck & Jones 2004). Therefore, a successful DNA
or subunit protein-based vaccine will likely require a cocktail of proven immunogens.
Accordingly, there is increasing emphasis on strategies for combining protective
antigen candidates in the same regimen (Campos-Neto et al. 2002, Skeiky et al.
2002, Iborra et al. 2004, Zadeh-Vakili et al. 2004, Moreno et al. 2007, Rodriguez-Cortés
et al. 2007, Zanin et al. 2007). It should be noted that a triple fusion protein
vaccine (termed Leish-111f-MPL®-SE), consisting of the T-cell
adjuvant antigens thiol-specific antioxidant, Leishmania major stress-inducible
protein 1 and Leishmania elongation initiation factor formulated in monophosphoryl
lipid A plus squalene, which confers protection in the mouse model against L.
major, Leishmania amazonensis (Coler & Reed 2005) and Leishmania
infantum infections (Coler et al. 2007) is now within reach. Whether prophylactic
immunization using this vaccine can achieve similar levels of immunity against
all parasite species that cause disease in genetically diverse human subjects
(who also may differ significantly in their nutritional status and previous
immunological experience) has yet to be determined.
Additionally, the
potential efficacy of the Leish-111f/GM-CSF adjuvant vaccine in a post-exposure
paradigm is currently being tested in cases of drug-refractory disease with
encouraging results (Badaró et al. 2006). On the other hand, the potential
for immunomodulatory factors of sandfly saliva to serve as vaccine targets to
prevent pathogen transmission (Titus et al. 2006) has received increased attention
by investigators. In this regard, two candidates are the Lutzomyia longipalpis
salivary gland protein maxadilan (Brodie et al. 2007) and the recombinant protein
SP15; a vaccine composed of the latter antigen confers protection in the mouse
model against L. major challenge infection (Valenzuela et al. 2001).
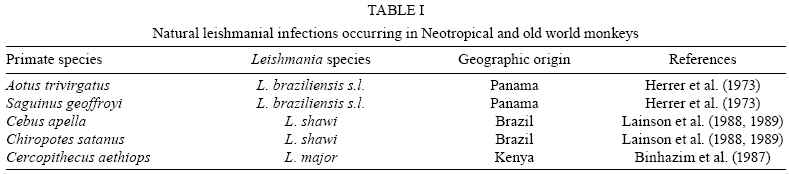
Natural and
experimental leishmanial infections in NHP
Table
I summarizes the published studies on natural leishmanial infections in
NHP. At least four species of Neotropical monkeys are susceptible to natural
infection with human pathogenic Leishmania (Viannia) species (Herrer
et al. 1973, Lainson et al. 1988, 1989). In contrast, only one species of old
world monkeys was found to be naturally infected with L. major (Binhazim
et al. 1987).
Monkeys have varying
degrees of susceptibility to leishmanial parasites and the specific disease
course depends on the challenge parasite (Amaral et al. 1996, 2001, Teva et
al. 2003), host species or individual (Dennis et al. 1986, Porrozzi et al. 2006)
challenge dose and route of exposure (Lujan et al. 1986a, Amaral et al. 1996).
Moreover, sand fly saliva immunomodulators are known to exacerbate leishmanial
infection in rodents (Lima & Titus 1996). Accordingly, when rhesus macaques
are infected with L. major transmitted by Phlebotomus papatasi
(Probst et al. 2001), they developed skin lesions that lasted longer (12-28
weeks post-infection) than typical infections (11 weeks) induced by needle inoculation
with larger numbers (1 x 107) of L. major culture metacyclics
(Amaral et al. 2001).
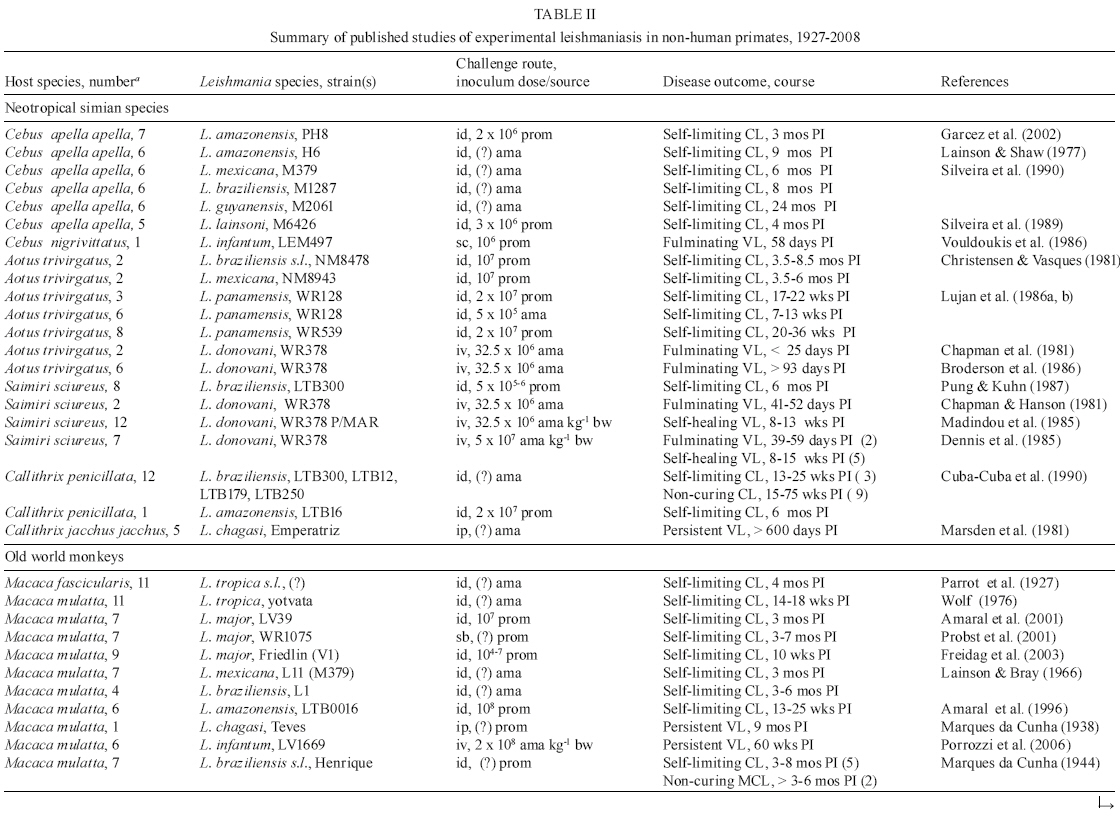
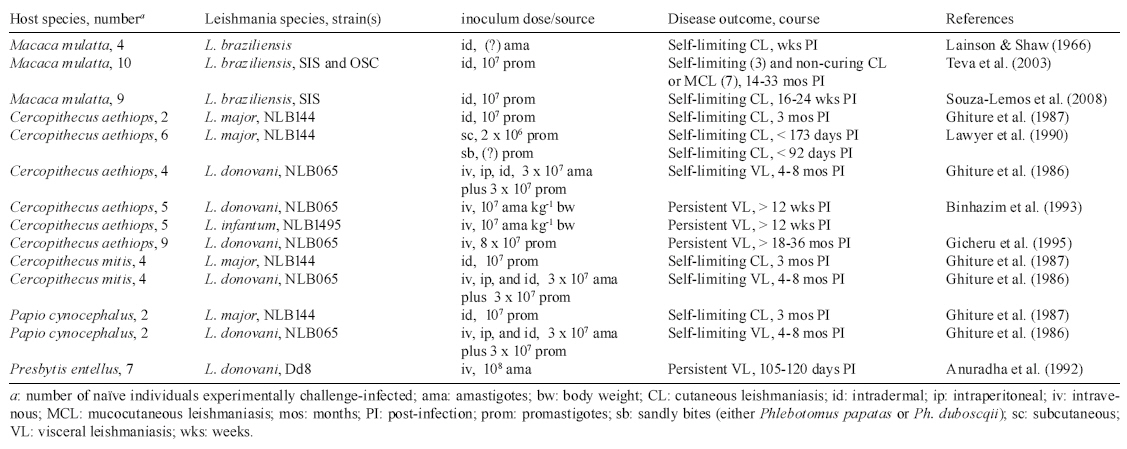
Table
II a, II b summarizes the essential features of the published studies on experimental
infection of NHP by various Leishmania species. Different NHP species
have become useful in studying the biology of infection and in dissecting the
host response to these parasites. Those reported as being highly susceptible
to Leishmania donovani complex parasites include the Neotropical simians
Aotus trivirgatus (Chapman et al. 1981, Broderson et al. 1986), Saimiri
sciureus (Chapman & Hanson 1981, Dennis et al. 1985, 1986) and Callithrix
jacchus jacchus (Marsden et al. 1981). All of these species have since been
used as NHP models of VL for anti-leishmanial chemotherapy studies (Dietze et
al. 1985, Madindou et al. 1985, Berman et al. 1986). Conversely, East African
primates such as Sykes monkeys (Cercopithecus mitis) and baboons (Papio
cynocephalus) all supported low-grade L. donovani infections for
periods ranging between 4-8 months and subsequently showed evidence of self-cure
(Githure et al. 1986). Furthermore, disease mimicking human VL was established
in langur monkeys (Presbytis entellus) (Dube et al. 1999), vervet monkeys
(Cercopithecus aethiops) (Binhazim et al. 1993, Gicheru et al. 1995)
and macaques (Macaca mulatta) (Porrozzi et al. 2006). The L. donovani-langur
monkey model has also been explored to assess different vaccine formulations
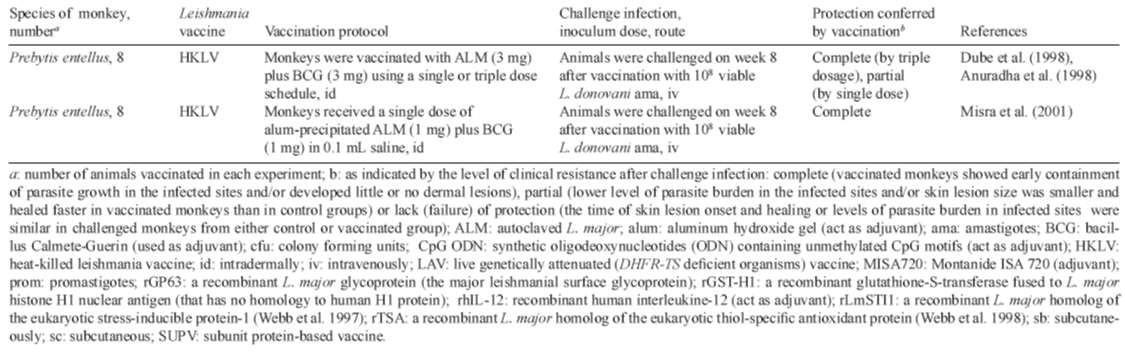
against VL (Dube et al. 1998, Misra et al. 2001).
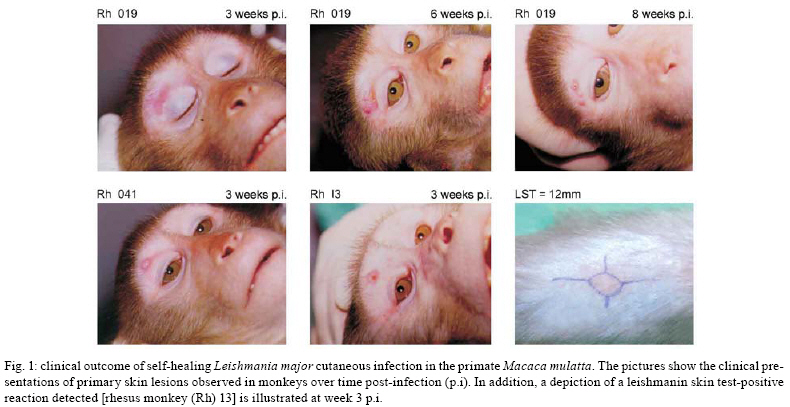
Consistent with
documented cases of human CL caused by L. major, experimental infection
in macaques induced by the same parasite species causes a self-limiting CL of
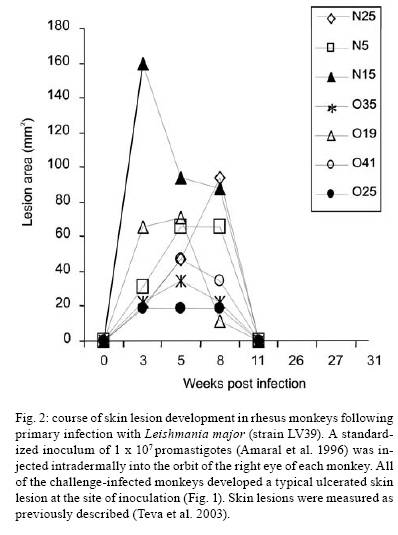
moderate severity (Fig. 1), which resolves within
three months (Fig. 2) and provides the most ethically acceptable
model for vaccine testing (Amaral et al. 2001, 2002, Campos-Neto et al. 2001).
When infected with L. amazonensis, macaques developed greater lesion
size with longer duration (Amaral et al. 1996). In both experiments, active
skin lesions contained amastigotes with a mononuclear infiltrate of macrophages,
plasma cells and lymphocytes and formation of tuberculoid-type granulomas. In
L. amazonensis-infected macaques it was demonstrated that CD4+/CD8+
T-cell ratios favour CD8+ cells in both active and healing skin lesions
(Amaral et al. 2000). A more marked variation in the clinical course of infection
was found when groups of macaques were inoculated with different Leishmania
braziliensis strains (Teva et al. 2003, Souza-Lemos et al. 2008). The inocula
produced lesions of variable severity, ranging from localized self-healing CL
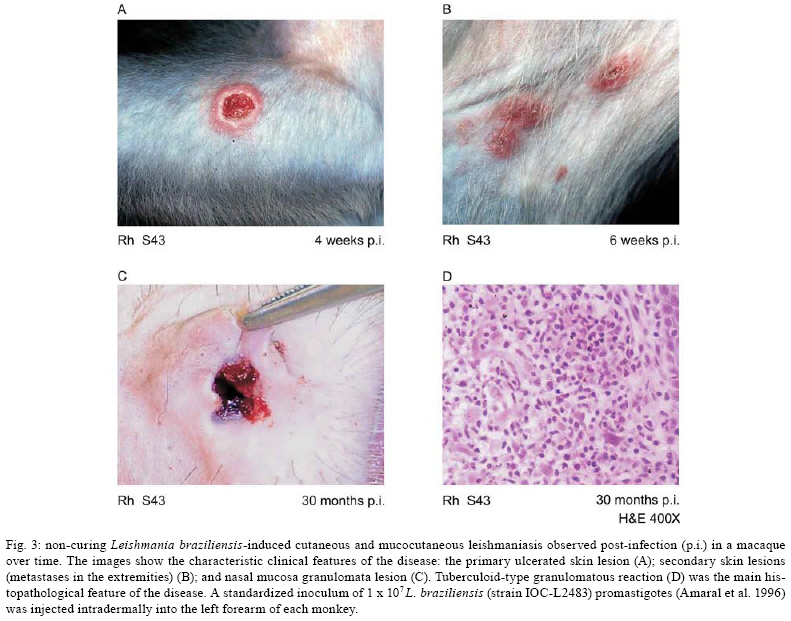
to non-healing disease (Figs 3A, C). Pathological findings included a typical
cell-mediated immunity-induced granulomatous reaction (Fig.
3D), which consisted of all cell types found within human granulomas, including
the presence of both IFN-γ- or TNF-α-producing
CD4+ and CD8+ T-cells, as well as IL-10-producing CD4+CD25+
T-cells (Souza-Lemos et al. 2008). While several groups have described that
ML (mucosa lesions) has not been observed in Neotropical monkey models of CL
(Lainson & Shaw 1977, Lujan et al. 1986a, 1990, Cuba Cuba et al. 1990),
in our own studies (Teva et al. 2003, G Grimaldi Jr, unpublished data) two of
30 (6.7%) L. braziliensis-infected macaques developed nasal ML (Fig.
3C). In the original model description (Marques da Cunha 1944), ML was observed
in two of seven (28.5%) monkeys infected with L. braziliensis. Of note,
therapeutic responses of L. braziliensis-infected macaques to the reference
drug N-methylglucamine antimoniate (Glucantime®) were
consistent with those reported in human disease (Teva et al. 2005).
Contrary to the
traditional belief that human self-resolution of CL confers life-long immunity
against further infection by the same parasite (Gunders 1987), Killick-Kendrick
et al. (1985) and Saraiva et al. (1990) provided evidence that immunity conferred
by prior self-resolving leishmanial infection may not always be complete in
humans. Likewise, in L. amazonensis (Amaral et al. 1996) or L. major-infected
out-bred macaques (Amaral et al. 2001) both the level of resistance and the
acquired immune response to subsequent homologous challenge(s) are variable.
The mechanism causing partial protection in primates is not yet clear, but may
be related to differential performance of memory T cells (Zaph et al. 2004).
In addition, IL-10-producing CD4+CD25+T cells are known
to control acquired immunity in mice (Belkaid et al. 2002) and macaques (Souza-Lemos
et al. 2008) with leishmanial infections.
The findings from
cross-immunity experiments between different species or strains of Leishmania
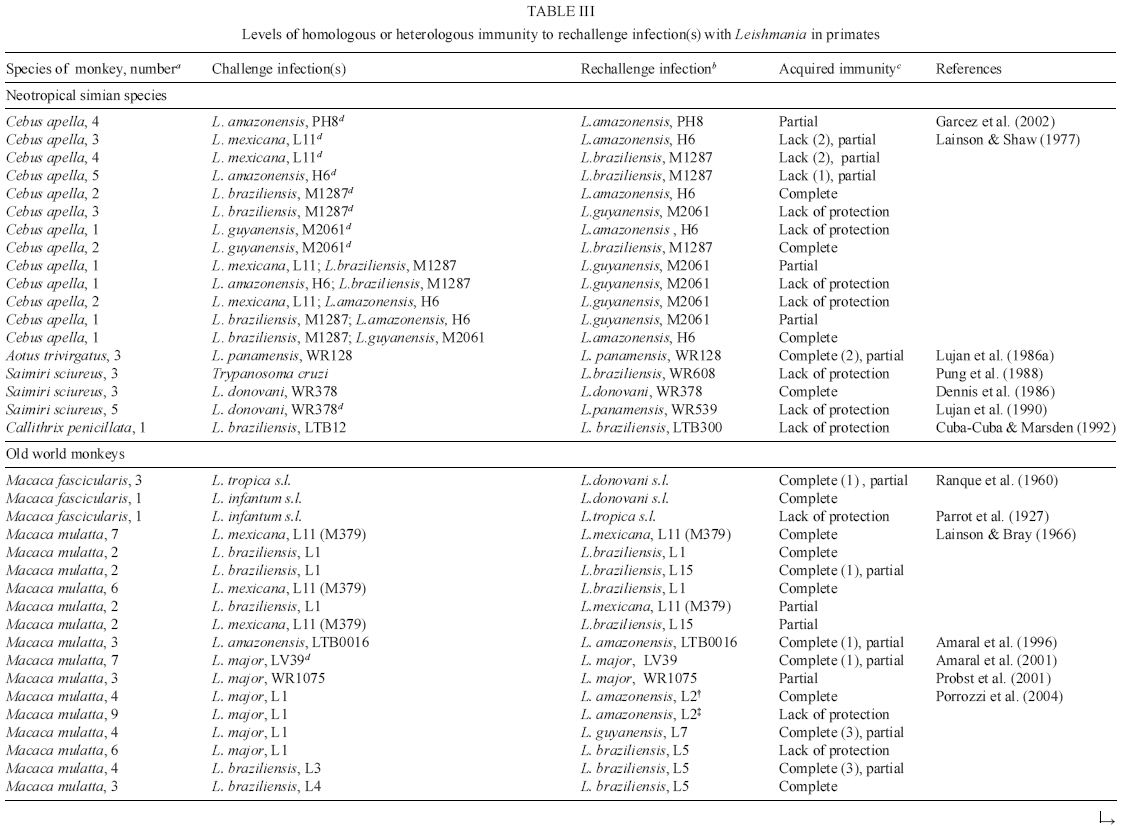
in monkeys (Table IIIa , IIIb ) may give important clues
to vaccine reseach. The relative variability in protection after self-cure or
drug-cured experimental leishmaniasis to challenge by heterologous parasites
appears to reflect both the nature (i.e., etiologic agent) and the course of
primary infection or disease tempo (i.e., the progression and resolution of
leishmanial lesions). Another factor that can influence acquired immunity is
the time between recovery from primary infection and re-challenge. For example,
a self-healing CL following infection with L. major induces significant
protection for L. amazo-nensis and Leishmania guyanensis and was
dependent on time of re-challenge by L. amazonensis after animals had
recovered from primary lesions, but lacked protection against L. braziliensis.
Conversely, macaques immune to either L. braziliensis or Leishmania
chagasi (syn. L. infantum) were found to be fully protected to challenge
with L. braziliensis or L. amazonensis (Porrozzi et al. 2004).
All infected animals
responded with increased production of immunoglobulins capable of binding to
cross-reacting parasite antigens (Lujan et al. 1987, Porrozzi et al. 2004).
Although an antigen-specific Th1-like response appears critical for mediating
protection in a variety of primate models of CL (Olobo et al. 1992, Olobo &
Reid 1993, Amaral et al. 2001, Teva et al. 2003) and VL (Porrozzi et al. 2006),
the current parameters of cell-mediated immunity [i.e., by measuring delayed-type
hypersensitivity reaction (DHT) to the leishmanin skin test (LST) in vitro lymphocyte
proliferation and IFN-γ production] do not always
correlate with clinical recovery and resistance to infectious re-challenge (Amaral
et al. 2001, Porrozzi et al. 2004, 2006). Certainly, further studying the immune
response in primates may clarify what is required to develop and maintain protective
immunity to re-challenge(s).
Use of primate
models to assess leishmaniasis vaccines
Divergent evolution
(~ 210 million year divergence between rodents and humans) limits the relevance
of murine models in guiding the design of human vaccine trials (Nikolich-Žugich
2007). In this regard, old world simian species which diverged from humans approximately
25 million years ago (Gibbs et al. 2007) are emerging as invaluable in vivo
models of pathogenesis and immunity to infectious diseases requiring cellular
immunity, but are also a key tool for conducting comparative studies of vaccine
approaches (Nathansson et al. 1999, Jonhston 2000). Because of the homology
between the M. mulatta and human immune systems (Kennedy et al. 1997b,
Shearer et al. 1999, Pahar et al. 2003, Giavedoni 2005), the NHP model is frequently
used to determine which vaccine candidates are most worthy of accelerated development
(Johnston 2000, Nikolich-Žugich 2007).
A variety of NHP
models for both CL and VL have been used to assess the safety (to verify whether
vaccination itself causes adverse effects), immunogenicity (including evaluation
of potential correlates of immune protection) and protective efficacy (to determine
whether vaccination protects the animal host against infective challenge) of
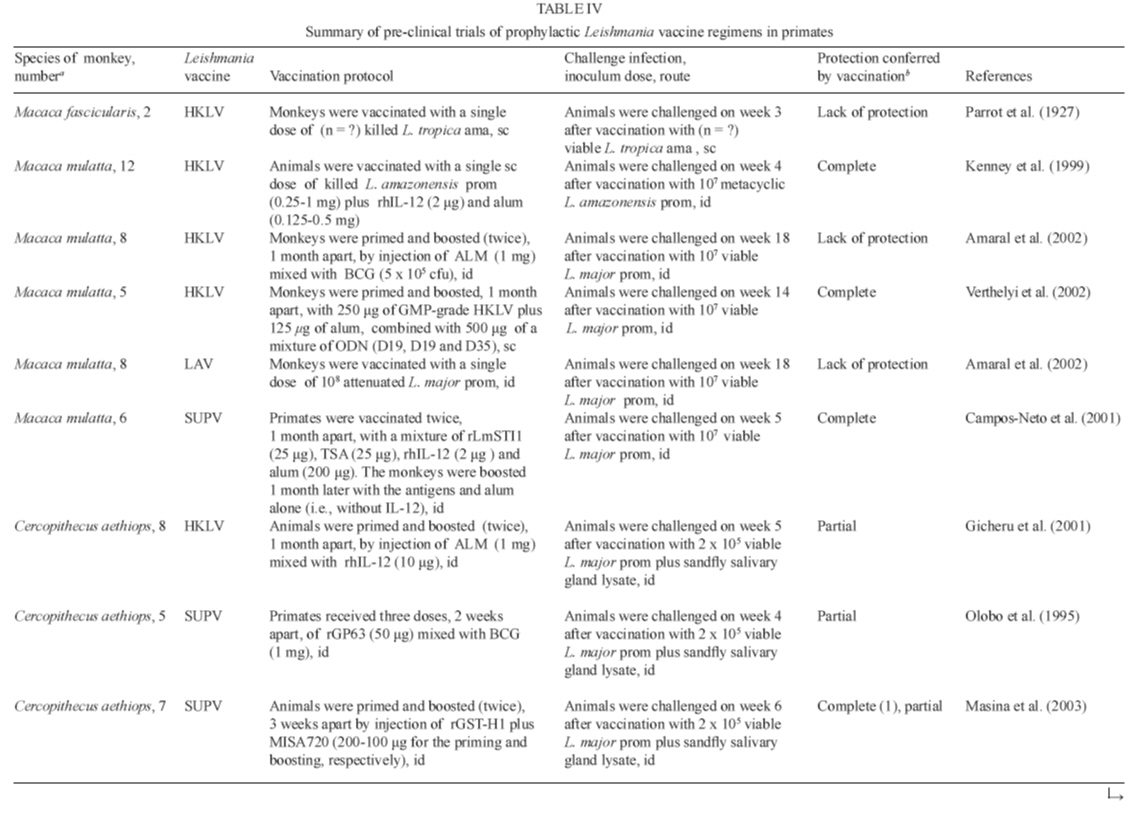
vaccine formulations (Table IV a, IV b). To date, the
only way to determine acquired resistance afforded by a candidate vaccine is
to challenge the vaccinated animals with virulent leishmanial parasites. However,
because of (i) the limited number of monkeys per experimental group and (ii)
the fact that stationary-phase promastigotes can have varying numbers of the
infectious form of metacyclic promastigotes within each preparation, researchers
use a high inoculum dose to achieve uniform infection for challenge, which may
account for the relative variability in the levels of vaccine-induced protection.
On the other hand, the use of a short interval between the last boost and the
infectious challenge (as short as 3-5 weeks in some studies), makes it difficult
to interpret the results in terms of the ability of the vaccine to induce a
sustained memory T-cell response (Pitcher et al. 2002). In addition, in most
studies of this nature, it is difficult to accurately assess partial host immunity
during infection since lesion size, a highly variable parameter (due to the
out-bred nature of monkeys used for such studies) is commonly used as a correlate
of protection.
The results from
primate vaccine studies are summarized in Table IV a, IV b.
Protective efficacy with crude vaccine approaches against CL in macaques was
achieved only when the inactivated parasites were combined with alum plus recombinant
human IL-12 (Kenney et al. 1999) or CpG ODN (Verthelyi et al. 2002) as adjuvants.
In addition, successful vaccination against L. donovani visceral infection
in langur monkeys was obtained using alum-precipitated autoclaved L. major
with BCG (Misra et al. 2001). In our previous studies (Amaral et al. 2002) we
have compared the potential efficacy of two L. major vaccines, one genetically
attenuated (DHFR-TS deficient organisms), the other inactivated organisms
(autoclaved promastigotes with BCG), in protecting macaques against homologous
challenge. While a positive antigen-specific recall proliferative response was
observed in those vaccinated (79% in attenuated parasite-vaccinated monkeys,
versus 75% in ALM-plus-BCG-vaccinated animals), none of these animals exhibited
either augmented in vitro INF-γ production or a positive
DTH response to the leishmanin skin test prior to challenge. Following challenge,
significant differences in blastogenic responses were found between attenuated-vaccinated
monkeys and naïve controls. Protective immunity did not follow vaccination,
in that monkeys exhibited skin lesions at the site of challenge in all experimental
groups. In contrast, vaccination using a mix of the recombinant antigens LmSTI1
and TSA (Webb et al. 1996, 1998) formulated with rhIL-12 and alum as adjuvants
induced excellent protection in the high dose L. major-macaque model
(Campos-Neto et al. 2001). Likewise, vervet monkeys, when immunized with recombinant
histone H1 antigen using Montanide as an adjuvant, mounted good protection against
challenge with L. major (Masina et al. 2003).
Ample evidence
supports the notion that different prime-boost vaccination regimens can elicit
greater immune responses than single immunization modalities. The use of heterologous
prime-boost approaches was originally explored as a means to overcome vector-specific
immunity elicited against the priming immunogen and to augment antigen-specific
responses by subunit protein boost (Hu et al. 1991). This approach was found
to enhance antigen-specific antibody responses in mice, macaques and humans
primed with a recombinant vaccinia virus and boosted with recombinant HIV-1
envelope protein (Hu 2005). Conversely, immunization with DNA priming and recombinant
virus boosting elicited strong T-cell responses (Schneider et al.1999, Barouch
& Letvin 2000). The effect regarding the order of DNA versus recombinant
vector for priming or boosting can have in eliciting protective immunity has
been debated (Hanke et al. 1998, McClure et al. 2000). Over the past three years,
several primate studies have been performed in our laboratory to establish vaccination
procedures, improve vaccine immunogenicity and minimize vector-specific immunity.
Indeed, it is now clear that detectable Leishmania-specific T-cell responses
can be induced safely in primates by vaccination, but, depending on the particular
regimen used, varying degrees of acquired immunity have been achieved (ranging
from non-existent to full protection after the infectious challenge). Further
experiments are in progress in the Leishmania-macaque model to comparatively
examine the potential efficacy of various vaccine approaches afforded by vaccine
candidates.
Determining
correlates of immune protection to Leishmania
While the functional
heterogeneity (across HLA haplotypes) of T-cell cytokine responses to existing
vaccines is undoubtedly of importance (Robinson & Amara 2005), they have
not been extensively analyzed. In fact, T-cell vaccines for microbial infections
have been developed without a clear understanding of their mechanism(s) of protection
(Lambert et al. 2005). With regard to leishmaniasis, most vaccine studies measure
the frequency of IFN-γ-producing Th1 cells as the
primary immune correlate of protection (Coller & Reed 2005), but in vitro
IFN- γ production as a single immune parameter may
not be sufficient to predict protection (Elias et al. 2005, Oliveira et al.
2005). Recent data have shed important insight on the potential correlates of
protection, showing that the magnitude, potency and durability of a multifunctional
CD4+ Th1-cell cytokine response can be a crucial determinant in whether
a vaccine is protective (Darrah et al. 2007). Conversely, it is likely that
IL-10-producing CD4+CD25+ T regulatory cells are functional
in determining vaccine failure (Stober et al. 2005). In another study (Stäger
et al. 2000), vaccine-induced protection, using the recombinant hydrophilic
acylated surface protein B1 (HASPB1) of L. donovani, correlates with
the presence of rHASPB1-specific, IFN-γ -producing
CD8+ T cells.
Neither study in
the L. amazonensis (Kenney et al. 1999) or L. major-macaque model
(Campos-Neto et al. 2001, Amaral et al. 2002), nor those in the L. major-vervet
monkey model (Gicheru et al. 2001), have resulted in a clear definition of what
T-cell responses will be required for vaccine-induced protective immunity. Without
such knowledge, vaccine design strategies will remain largely empirical, and
further failures are likely to occur. This is due, in part, to the high degree
of variability in the antigen-specific recall blastogenic and IFN-γ
responses detected among primates (Campos-Neto et al. 2001, Pahar et al. 2003).
This appears to result from the outbred genetics of macaques used for such studies,
which indeed makes them the most appropriate model when predicting the diversity
of responses that could be expected in the human population. Increasing the
number of monkeys per experimental group can help address this condition. Unfortunately,
by definition this is not feasible. On the other hand, using macaques with defined
genotypes with respect to immune response genes (MHC class I and II) would minimize
individual variability, but unfortunately this approach introduces bias into
the results (Johnston 2000, Hu 2005).
Finally, the application
of ELIspot and cytokine flow cytometry assays has provided new insights into
the attributes of both CD4+ and CD8+ T cells that mediate
protection in macaques (Mäkitalo et al. 2002, Keeney et al. 2003). This
technology should help to identify correlates of protection in future primate
vaccine studies.
Concluding remarks
Clinical development
of the available subunit protein or DNA-based vaccines against leishmaniasis
may not be fully protective across all HLA haplotypes and Leishmania
species. This is due, in part, to the inherent difficulties that hinder full
characterization of the determinants of successful T-cell immunity in humans
(Robinson & Amara 2005, Appay et al. 2008). Nevertheless, most experts believe
that a successful Leishmania vaccine will likely require (i) selection
of a cocktail of protective immunogens; (ii) identification of efficient prime-boost
strategies in order to provide broad, cross-reactive and long-lasting protection;
and (iii) selection or identification of an adjuvant formulations or delivery
systems that can be used in human clinical trials. Nonetheless, given these
intrinsic vaccine development requirements, regulatory authorities are willing
to regulate safety data on infectious vectored vaccines generated from primates.
However, primate
testing should be reserved for the final stages of evaluation of vaccine candidates
that have already shown consistent induction of significant protective immunity
in conventional mouse models. Considerations for employing the primate M.
mulatta to evaluate vaccine safety and protective efficacy should also include
costs and availability (Kennedy et al. 1997a). Available data indicates that
vaccine trials in macaques will not be hindered due to divergence of MHC class
I and class II molecules (Watkins et al. 1988, Klein et al. 1993, Doxiadis et
al. 2001). Moreover, rhesus macaques have been successfully infected with a
variety of human pathogenic Leishmania species either by syringe or sandfly
challenge and develop a human-like disease (including the non-curing L. braziliensis
granulomata ML). Most of the published information on the course of primary
or secondary infection, clinicopathological changes, immune responses and vaccination
data was gained using outbred macaques. Although the predictive value for any
animal model in vaccine development ultimately depends on validating data from
human trials, further development of the Leishmania-macaque model should
prove useful in guiding the design of human vaccine trials.
ACKNOWLEDGEMENT
To Dr. Antonio
Campos-Neto, for critically reviewing this manuscript.
REFERENCES
- Amaral VF, Pirmez
C, Ferreira AJS, Ferreira V, Grimaldi Jr G 2000. Cell populations in lesions
of cutaneous leishmaniasis of Leishmania (L.) amazonensis-infected
rhesus macaques, Macaca mulatta. Mem Inst Oswaldo Cruz 95: 209-216.
- Amaral VF, Ransatto
VAO, Conceição-Silva F, Molinaro E, Ferreira V, Coutinho SG, McMahon-Pratt
D, Grimaldi G Jr 1996. Leishmania amazonensis: the Asian rhesus macaques
(Macaca mulatta) as an experimental model for study of cutaneous leishmaniasis. Exp Parasitol 82: 34-44.
- Amaral VF, Teva
A, Oliveira-Neto MP, Silva AJ, Pereira MS, Cupolillo E, Porrozzi R, Coutinho
SG, Pirmez C, Beverley SM, Grimaldi G Jr 2002. Study of the safety, immunogenicity
and efficacy of attenuated and killed Leishmania (Leishmania) major vaccines
in a rhesus monkey (Macaca mulatta) model of the human disease. Mem
Inst Oswaldo Cruz 97: 1041-1048.
- Amaral VF, Teva
A, Porrozzi R, Silva AJ, Pereira MS, Grimaldi G Jr 2001. Leishmania (Leishmania) major-infected rhesus macaques (Macaca mulatta) develop varying
levels of resistance against homologous reinfections. Mem Inst Oswaldo Cruz
96: 795-804.
- Anuradha R, Pal
R, Zehra K, Katiyar JC, Sethi N, Bhatia G, Singh RK 1992. The Indian langur:
preliminary report of a new non-human primate host for visceral leishmaniasis. WHO Bulletin 70: 63-72.
- Anuradha D, Sharma
P, Skrivastava JK, Misra A, Naik S 1998. Vaccination of langur monkey (Presbytis
entellus) against Leishmania donovani with autoclaved L. major plus BCG. Parasitology 116: 219-221.
- Appay V, Douek
DC, Price DA 2008. CD8+ T cell efficacy in vaccination and disease. Nature Med 14: 623-628.
- Badaró R,
Lobo I, Munos A, Netto EM, Modabber F, Campos-Neto A, Coler RN, Reed SG 2006.
Immunotherapy for drug-refractory mucosal leishmaniasis. J Infect Dis 194:
1151-1159.
- Barouch DH, Letvin
NL 2000. DNA Vaccination for HIV-1 and SIV. Intervirology 43: 282-287.
- Belkaid Y, Piccirillo
CA, Mendez S, Shevach EM, Sacks DL 2002. CD4+CD25+ regulatory
T cells control Leishmania major persistence and immunity. Nature
420: 502-507.
- Berman JD, Hanson
WL, Chapman WL, Alving CR, Lopez-Berestein G 1986. Antileishmanial activity
of liposome-encapsulated Amphotericin B in hamsters and monkeys. Antimicrob
Agents Chemother 30: 847-851.
- Bhowmick S, Ravindran
R, Ali N 2007. Leishmanial antigens in liposomes promote protective immunity
and provide immunotherapy against visceral leishmaniasis via polarized Th1 response. Vaccine 25: 6544-6556.
- Binhazim AA, Githure
JI, Muchemi GK, Reid GD 1987. Isolation of Leishmania major from a naturally
infected vervet monkey (Cercopithecus aethiops) caught in Kiambu District,
Kenya. Parasitology 73: 1278-1279.
- Binhazim AA, Shin
SS, Chapman WL Jr, Olobo J 1993. Comparative susceptibility of African green
monkeys (Cercopithecus aethiops) to experimental infection with Leishmania
leishmania donovani and Leishmania leishmania infantum. Lab Anim
Sci 43: 37-47.
- Broderson JR, Chapman
WL Jr, Hanson WL 1986. Experimental visceral leishmaniasis in the owl monkey. Vet Pathol 23: 293-302.
- Brodie TM, Smith
MC, Morris RV, Titus RG 2007. Immunomodulatory effects of the Lutzomyia longipalpis salivary gland protein Maxadilan on mouse macrophages. Infect Immun 75:
2359-2365.
- Campos-Neto A,
Porrozzi R, Greeson K, Coler RN, Webb JR, Seiky YA, Reed SG, Grimaldi G Jr 2001.
Protection against cutaneous leishmaniasis induced by recombinant antigens in
murine and non-human primate models of the human disease. Infect Immun 69:
4103-4108.
- Campos-Neto A,
Webb JR, Greeson K, Coler RN, Skeiky YA, Reed SG 2002. Vaccination with plasmid
DNA encoding TSA/LmSTI1 leishmanial fusion proteins confers protection against Leishmania major infection in susceptible BALB/c mice. Infect Immun
70: 2828-2836.
- Chapman WL Jr,
Hanson WL 1981. Visceral leishmaniasis in the squirrel monkey (Saimiri sciurea). J Parasitol 67: 740-741.
- Chapman WL Jr,
Hanson WL, Hendricks LD 1981. Leishmania donovani in the owl monkey (Aotus
trivirgatus). Trans R Soc Trop Med Hyg 75: 124-125.
- Christensen HA,
de Vasquez AM 1981. Susceptibility of Aotus trivirgatus to Leishmania
braziliensis and L. mexicana. Am J Trop Med Hyg 30: 54-56.
- Coler RN, Goto
Y, Bogatzki L, Raman V, Reed SG 2007. Leish-111f, a recombinant polyprotein
vaccine that protects against visceral leishmaniasis by elicitation of CD4+ T cells. Infect Immun 75: 4648-4654.
- Coler RN, Reed
SG 2005. Second-generation vaccines against leishmaniasis. Trends Parasitol
21: 244-248.
- Cuba-Cuba CA, Ferreira
V, Bampi M, Magalhães A, Marsden P, Vexenat A, Mello MT 1990. Experimental
infection with Leishmania (Viannia) braziliensis and Leishmania (Leishmania)
amazonensis in marmoset (Callithrix penicillata) (Primates, Callithricidae). Mem Inst Oswaldo Cruz 85: 459-467.
- Cuba-Cuba CA, Marsden
PD 1992. Failure to develop homologous immunity to a second challenge with Leishmania
(Viannia) braziliensis in the black-plumed marmoset (Callithrix penicillata). Trans R Soc Trop Med Hyg 86: 37-45.
- Darrah PA, Patel
DT, De Luca PM, Lindsay RWB, Davey DF, Flynn BJ, Hoff ST, Andersen P, Reed SG,
Morris SL, Roederer M, Seder RA 2007. Multifunctional TH1 cells define
a correlate of vaccine-mediated protection against Leishmania major.
Nature Med 13: 843-850.
- Davies CR, Kaye
P, Croft SL, Sundar S 2003. Leishmaniasis: new approaches to disease control. BMJ 326: 377-382.
- Delgado G, Parra-López
C, Spinel C, Patarroyo ME 2005. Phenotypical and functional characterization
of non-human primate Aotus spp. dendritic cells and their use as a tool
for characterizing immune response to protein antigens. Vaccine 23: 3386-3395.
- Dennis VA, Chapman
WL Jr, Hanson WL, Lujan R 1985. Leishmania donovani: clinical, haematological
and hepatic changes in squirrel monkeys (Saimiri sciureus). J Parasitol
71: 576-582.
- Dennis VA, Lujan
R, Chapman WL Jr, Hanson WL 1986. Leishmania donovani: Cellular and humoral
immune responses after primary and challenge infections in squirrel monkeys, Saimiri sciureus. Exp Parasitol 61: 319-334.
- Dietze R, Araújo
RC, Lima MLR, Vexenat JA, Marsden PD, Barreto AC 1985. Ensaio terapêutico
com glucantime em sagüis (Callithrix jacchus) infectados com uma
cepa de Leishmania donovani aparentemente resistente ao tratamento. Rev
Soc Bras Med Trop 18: 39-42.
- Dondji B, Deak
E, Goldsmith-Pestana K, Perez-Jimenez E, Esteban M, Miyake S, Yamamura T, McMahon-Pratt
D 2008. Intradermal NKT cell activation during DNA priming in heterologous prime-boost
vaccination enhances T cell responses and protection against Leishmania. Eur J Immunol 38: 706-719.
- Dondji B, Pérez-Jimenez
E, Goldsmith-Pestana K, Esteban M, McMahon-Pratt D 2005. Heterologous prime-boost
vaccination with the LACK antigen protects against murine visceral leishmaniasis. Infect Immun 73: 5286-5289.
- Doxiadis GGM, Otting
NG, Natasja G, Bontrop RE 2001. Differential evolutionary MHC class II strategies
in humans and rhesus macaques: relevance for biomedical studies. Non-human primate
models for human disease and immunobiology. Immunol Rev 183: 76-85.
- Dube A, Sharma
P, Srivastava JK, Misra A, Naik S, Katiyar JC 1998. Vaccination of langur monkeys
(Presbytis entellus) against Leishmania donovani with autoclaved L. major plus BCG. Parasitology 116: 219-221.
- Dube A, Srivastava
JK, Sharma P, Chaturvedi A, Katiyar JC, Naik S 1999. Leishmania donovani:
cellular and humoral immune responses in Indian langur monkeys, Presbytis
entellus. Acta Trop 73: 37-48.
- Elias D, Akuffo
H, Britton S 2005. PPD induced in vitro interferon gamma production is
not a reliable correlate of protection against Mycobacterium tuberculosis. Trans R Soc Trop Med Hyg 99: 363-368.
- Freidag BL, Mendez
S, Cheever AW, Kenney RT, Flynn B, Sacks DL, Seder RA 2003. Immunological and
pathological evaluation of rhesus macaques infected with Leishmania major. Exp Parasitol 103: 160-168.
- Garcez LM, Silveira
FT, El Harith A, Lainson R, Shaw JJ 2002. Experimental cutaneous leishmaniasis
IV. The humoral response of Cebus apella (Primates: Cebidae) to infections
of Leishmania (Leishmania) amazonensis, L. (Viannia) lainsoni and L. (V.) braziliensis using the direct agglutination test. Acta
Trop 68: 65-76.
- Giavedoni LD 2005.
Simultaneous detection of multiple cytokines and chemokines from nonhuman primates
using luminex technology. J Immunol Methods 301: 89-101.
- Gibbs RA, Rogers
J, Katze MG, Bumgarner R, Weinstock GM, Mardis ER, Remington KA, Strausberg
RL, Venter JC, Wilson RK, Batzer MA, Bustamante CD, Eichler EE, Hahn MW, Hardison
RC, Makova KD, Miller W, Milosavljevic A, Palermo RE, Siepel A, Sikela JM, Attaway
T, Bell S, Bernard KE, Buhay CJ, Chandrabose MN, Clay Davis DM, Delehaunty KD,
Ding Y, Dinh HH, Dugan-Rocha S, Fulton LA, Gabisi RA, Garner TT, Godfrey J,
Hawes AC, Hernandez J, Hines S, Holder M, Hume J, Jhangiani SN, Joshi V, Khan
ZM, Kirkness EF, Cree A, Fowler RG, Lee S, Lewis LR, Li Z, Liu Y, Moore SM,
Muzny D, Nazareth LV, Ngo DN, Okwuonu GO, Pai G, Parker D, Paul HA, Pfannkoch
C, Pohl CS, Rogers Y-H, Ruiz SJ, Sabo A, Santibanez J, Schneider BW, Smith SM,
Sodergren E, Svatek AF, Utterback TR, Vattathil S, Warren W, White CS, Chinwalla
AT, Feng Y, Halpern AL, Hillier LW, Huang X, Minx P, Nelson JO, Pepin KH, Qin
X, Sutton GG, Venter E, Walenz BP, Wallis JW, Worley KC, Yang S-P, Jones SM,
Marra MA, Rocchi M, Schein JE, Baertsch R, Clarke L, Csürös M, Glasscock
J, Harris RA, Havlak P, Jackson AR, Jiang H, Liu Y, Messina DN, Shen Y, Xing-Zhi
Song H, Wylie T, Zhang L, Birney E, Han K, Konkel MK, Lee J, Smit AFA, Ullmer
B, Wang H, Xing J, Burhans R, Cheng Z, Karro JE, Ma J, Raney B, She X, Cox MJ,
Demuth JP, Dumas LJ, Han S-G, Hopkins J, Karimpour-Fard A, Kim YH, Pollack JR,
Vinar T, Addo-Quaye C, Degenhardt J, Denby A, Hubisz MJ, Indap A, Kosiol C,
Lahn BT, Lawson HA, Marklein A, Nielsen R, Vallender EJ, Clark AG, Ferguson
B, Hernandez RD, Hirani K, Kehrer-Sawatzki H, Kolb J, Patil S, Pu L-L, Ren Y,
Smith DG, Wheeler DA, Schenck I, Ball EV, Chen R, Cooper DN, Giardine B, Hsu
F, Kent FW, Lesk A, Nelson DL, O'Brien WE, Prüfer K, Stenson PD, Wallace
JC, Ke H, Liu X-M, Wang P, Xiang AP, Yang F, Barber GP, Haussler D, Karolchik
D, Kern AD, Kuhn RM, Smith KE, Zwieg AS 2007. Rhesus macaque genome sequencing
and analysis consortium. Evolutionary and biomedical insights from the rhesus
macaque genome. Science 316: 222-234.
- Gicheru MM, Olobo
JO, Anjili CO 1997. Heterologous protection by Leishmania donovani for Leishmania major infections in the vervet monkey model of the disease. Exp Parasitol 85: 109-116.
- Gicheru MM, Olobo
JO, Anjili CO, Orago AS, Modabber F, Scott P 2001. Vervet monkeys vaccinated
with killed Leishmania major parasites and interleukin-12 develop a type
1 immune response but are not protected against challenge infection. Infect
Immun 69: 245-251.
- Gicheru MM, Olobo
JO, Kariuki TM, Adhiambo C 1995. Visceral leishmaniasis in vervet monkeys: immunological
responses during asymptomatic infections. Scand J Immunol 41: 202-208.
- Githure JI, Reid
GD, Binhazim AA, Anjili CO, Shatry AM, Hendricks LD 1987. Leishmania major:
the suitability of East African nonhuman primates as animal models for cutaneous
leishmaniasis. Exp Parasitol 64: 438-447.
- Githure JI, Shatry
AM, Tarara R, Chulay JD, Suleman MA, Chunge CN, Else JG 1986. The suitability
of East African primates as animal models of visceral leishmaniasis. Trans
R Soc Trop Med Hyg 80: 575-576.
- Gonzalo RM, del
Real G, Rodriguez JR, Rodriguez D, Heljasvaara R, Lucas P, Larraga V, Esteban
M 2002. A heterologous prime-boost regime using DNA and recombinant vaccinia
virus expressing the Leishmania infantum P36/LACK antigen protects BALB/c
mice from cutaneous leishmaniasis. Vaccine 20: 1226-1231.
- Grimaldi G Jr 1995.
Meeting on vaccine studies towards the control of leishmaniasis. Mem Inst
Oswaldo Cruz 90: 553-556.
- Grimaldi G Jr,
Tesh RB 1993. Leishmaniasis of the New World: Current concepts and implications
for future research. Clin Microbiol Rev 6: 230-250.
- Gunders AE 1987.
Vaccination: past and future role in control. In W Peters, R Killick-Kendrick
(eds.), The leishmaniasis in Biology and Medicine, Vol. 2, Academic Press,
London, p. 928-941.
- Gurunathan S, Stobie
L, Prussin C, Sacks DL, Glaichenhaus N, Fowell DJ, Locksley RM, Chang JT, Wu
C-Y, Seder RA 2000. Requirements for the maintenance of Th1 immunity in vivo following DNA vaccination: a potential immunoregulatory role for CD8+ T cells. J Immunol 165: 915-924.
- Hanke T, Blanchard
TJ, Schneider J, Hannan CM, Becker M, Gilbert SC, Hill AVS, Smith GL, McMichael
A 1998. Enhancement of MHC class I-restricted peptide-specific T cell induction
by a DNA prime/MVA boost vaccination regime. Vaccine 16: 439-445.
- Hein WR, Griebel
PJ 2003. A road less traveled: large animal models in immunological research. Nat Rev Immunol 3: 79-84.
- Herrer A, Christensen
HA, Beumer RJ 1973. Reservoir hosts of cutaneous leishmaniasis among Panamanian
forest mammals. Am J Trop Med Hyg 22: 585-591.
- Hu S-L 2005. Non-human
primate models for AIDS vaccine research. Curr Drug Targets Infect Disord
5: 193-201.
- Hu S-L, Klaniecki
J, Dykers T, Sridhar P, Travis BM 1991. Neutralizing antibodies against HIV-1
BRU and SF2 isolates generated in mice immunized with recombinant vaccinia virus
expressing HIV-1 (BRU) envelope glycoproteins and boosted with homologous gp160. AIDS Res Hum Retroviruses 7: 615-620.
- Iborra S, Carrión
J, Anderson C, Alonso C, Sacks D, Soto M 2005. Vaccination with the Leishmania
infantum acidic ribosomal P0 protein plus CpG oligodeoxynucleotides induces
protection against cutaneous leishmaniasis in C57BL/6 mice but does not prevent
progressive disease in BALB/c mice. Infect Immun 73: 5842-5852.
- Iborra S, Soto
M, Carrión J, Alonso C, Requena JM 2004. Vaccination with a plasmid DNA
cocktail encoding the nucleosomal histones of Leishmania confers protection
against murine cutaneous leishmaniosis. Vaccine 22: 3865-3876.
- Johnston MI 2000.
The role of non-human primate models in AIDS vaccine development. Mol Med
Today 6: 267-270.
- Kedzierski L, Zhu
Y, Handman E 2006. Leishmania vaccines: progress and problems. Parasitology
133 (Suppl.): 87-112.
- Keeney TS, Nomura
LE, Maecker HT, Sastry KJ 2003. Flow cytometric analysis of macaque whole blood
for antigen-specific intracellular cytokine production by T lymphocytes. J
Med Primatol 32: 23-30.
- Kennedy RC, Shearer
MH, Hildebrand WH 1997a. Nonhuman primate models to evaluate vaccine safety
and immunogenicity. Vaccine 15: 903-908.
- Kennedy RC, Shearer
MH, Hildebrand WH, Simmonds RS 1997b. Non-human primates and their use in immunologically
based investigations. The Immunologist 5/5: 150-156.
- Kenney RT, Sacks
DL, Sypek JP, Vilela L, Gam AA, Evans-Davis K 1999. Protective immunity using
recombinant human IL-12 and alum as adjuvants in a primate model of cutaneous
leishmaniasis. J Immunol 163: 4481-4488.
- Killick-Kendrick
R, Bryceson ADM, Peter W, Evans DA, Leaney AJ, Rioux JA 1985. Zoonotic cutaneous
leishmaniasis in Saudi Arabia: Lesions healing naturally in man followed by
a second infection with the same zymodeme of Leishmania major. Trans
R Soc Trop Med Hyg 79: 363-365.
- Klein J, Satta
Y, O'Huigin C, Takahata N 1993. The molecular descent of the major histocompatibility
complex. Immunol Reviews 11: 213-244.
- Kostense S, Vandenberghe
K, Joling J, Van Baarle D, Nanlohy N, Manting E, Miedema F 2002. Persistent
numbers of tetramer+ CD8+ T cells, but loss of interferon-+ HIV-specific T cells during progression to AIDS. Blood 99: 2505-2511.
- Lainson R, Braga RR,
de Souza AAA, Povoa MM, Ishikawa EAY, Silveira FT 1989. Leishmania
(Viannia) shawi sp. n., a parasite of monkeys, sloths and procyo-nids in
Amazonian Brazil. Ann Parasitol Hum Comp 64: 200-207.
- Lainson R, Bray
RS 1966. Studies on the immunology and serology of leishmaniasis. II. Cross-immunity
experiments among different forms of American cutaneous leishmaniasis in monkeys. Trans R Soc Trop Med Hyg 60: 526-532.
- Lainson R, Shaw
JJ 1966. Studies on the immunology and serology of leishmaniasis. III. Cross-immunity
between Panamenian cutaneous leishmaniasis and Leishmania mexicana infection
in man. Trans R Soc Trop Med Hyg 60: 533-535.
- Lainson R, Shaw
JJ 1977. Leishmaniasis in Brazil: XII. Observations on cross-immunity in monkeys
and man infected with Leishmania mexicana mexicana, L. m. amazonensis, L. braziliensis braziliensis, L. b. guyanensis and L. b. panamensis. J Trop Med 80: 29-35.
- Lainson R, Shaw
JJ, Braga RR, Sacawa EAY, Souza AA, Silveira FT 1988. Isolation of Leishmania from monkeys in the Amazon region of Brazil. Trans R Soc Trop Med Hyg 82:
132.
- Lambert P-H, Liu
M, Siegrist C-A 2005. Can successful vaccines teach us how to induce efficient
protective immune responses? Nature Med 11 (Suppl.): 54-62.
- Lawyer PG, Ghiture
JI, Anjili CO, Olobo JO, Koech DK, Reid GDF 1990. Experimental transmission
of Leishmania major to vervet monkeys (Cercopithecus aethiops)
by bites of Phlebotomus duboscqi (Diptera: Psychodidae). Trans R Soc
Trop Med Hyg 84: 229-232.
- Levinson G, Hughes
AL, Letvin NL 1992. Sequence and diversity of rhesus monkey T-cell receptor
beta chain genes. Immunogenetics 35: 75-88.
- Lima HC, Titus
RG 1996. Effects of sand fly vector saliva on development of cutaneous lesions
and the immune response to Leishmania braziliensis in BALB/c mice. Infect
Immun 64: 5442-5445.
- Lujan R, Chapman
WL Jr, Hanson WL, Dennis VA 1986a. Leishmania braziliensis: development
of primary and satellite lesions in the experimentally infected owl monkey, Aotus trivirgatus. Exp Parasitol 61: 348-358.
- Lujan R, Chapman
WL Jr, Hanson WL, Dennis VA 1990. Leishmania braziliensis in the squirrel
monkeys: development of primary and satellite lesions and lack of cross-immunity
with Leishmania donovani. J Parasitol 76: 594-597.
- Lujan R, Dennis
VA, Chapman WL Jr, Hanson WL 1986b. Blastogenic responses of peripheral blood
leukocytes from owl monkeys experimentally infected with Leishmania braziliensis
panamensis. Am J Trop Med Hyg 35: 1103-1109.
- Lujan R, Hanson
WL, Chapman WL Jr, Dennis VA 1987. Antibody responses, as measured by the enzyme-linked
immunosorbent assay (ELISA), in owl monkeys experimentally infected with Leishmania
braziliensis panamensis. J Parasitol 73: 430-432.
- MacGregor RR, Boyer
JD, Ugen KE, Lacy KE, Gluckman SJ, Bagarazzi ML, Chattergoon MA, Baine Y, Higgins
TJ, Ciccarelli RB, Coney LR, Ginsberg RS, Weiner DB 1998. First human trial
of a DNA-based vaccine for treatment of human immunodeficiency virus type 1
infection: safety and host response. J Infect Dis 178: 92-100.
- Madindou TJ, Hanson
WL, Chapman WL Jr 1985. Chemotherapy of visceral leishmaniasis (Leishmania
donovani) in the squirrel monkey (Saimiri sciureus). Ann Trop
Med Parasitol 79: 13-19.
- Mäkitalo B,
Andersson M, Areström I, Karlén K, Villinger F, Ansari A, Paulie
S, Throstensson R, Ahlborg N 2002. ELISpot and ELISA analysis of spontaneous,
mitogen-induced and antigen-specific cytokine production in cynomolgus and rhesus
macaques. J Immunol Methods 270: 85-97.
- Marques da Cunha
A 1938. Infecções experimentais na leishmaniose visceral americana. Mem Inst Oswaldo Cruz 33: 581-598.
- Marques da Cunha
A 1944. Infecções experimentais na leishmaniose tegumentar americana. Mem Inst Oswaldo Cruz 41: 263-282.
- Marsden PD, Cuba
CC, Vexenat A, Costa e Silva M, Costa e Silva A, Barreto AC 1981. Experimental Leishmania chagasi infections in the marmoset Callithrix jacchus jacchus. Trans R Soc Trop Med Hyg 75: 314-315.
- Massina S, Gicheru
MM, Dempts SO, Fasel NJ 2003. Protection against cutaneous leishmaniasis in
outbred vervet monkeys, using a recombinant histone H1 antigen. J Infect
Dis 188: 1250-1257.
- McClure J, Schmidt
AM, Rey-Cuille M-A, Bannink J, Misher L, Tsai C-C, Anderson DM, Morton WR, Hu
S-L 2000. Derivation and characterization of a highly pathogenic isolate of
human immunodeficiency virus type 2 that causes rapid CD4+ cell depletion
in Macaca nemestrina. J Med Primatol 29: 114-126.
- Méndez S,
Gurunathan S, Kamhawi S, Belkaid Y, Moga MA, Skeiky YA, Campos-Neto A, Reed
S, Seder RA, Sacks D 2001. The potency and durability of DNA- and protein-based
vaccines against Leishmania major evaluated using low-dose, intradermal
challenge. J Immunol 166: 5122-5128.
- Misra A, Dube A,
Srivastava B, Sharma P, Srivastava JK, Katiyar JC, Naik S 2001. Successful vaccination
against Leishmania donovani infection in Indian langur using alum-precipitated
autoclaved Leishmania major with BCG. Vaccine 19: 3485-3492.
- Moreno J, Nieto
J, Masina S, Cañavate C, Cruz I, Chicharro C, Carrillo E, Napp S, Reymond
C, Kaye PM, Smith DF, Fasel N, Alvar J 2007. Immunization with H1, HASPB1 and
MML Leishmania proteins in a vaccine trial against experimental canine
leishmaniasis. Vaccine 25: 5290-5300.
- Nathansson N, Hirsch
VM, Mathieson BJ 1999. The role of non-human primates in thedevelopment of an
AIDS vaccine. AIDS 13 (Suppl. A): 113-120.
- Nikolich-Žugich
J 2007. Non-human primates models of T-cell reconstitution. Semin Immunol
19: 310-317.
- Oliveira MR, Tafuri
WL, Afonso LCC, Oliveira MAP, Nicoli JR, Vieira EC, Scott P, Melo MN, Vieira
LQ 2005. Germ-free mice produce high levels of interferon-gamma in response
to infection with Leishmania major but fail to heal lesions. Parasitology
131: 477-488.
- Olobo JO, Anjili
CO, Gicheru MM, Mbati PA, Kariuki TM, Githure JI, Koech DK, McMaster 1995. Vaccination
of vervet monkeys against cutaneous leishmaniasis using recombinant Leishmania "major surface glycoporotein" (gp63). Vet Parasitol 60: 199-212.
- Olobo JO, Reid
GDF 1993. Delayed-type hypersensitivity responses in vervet monkeys self-cured
from experimental cutaneous leishmaniasis. Acta Trop 52: 309-311.
- Olobo JO, Reid
GDF, Githure JI, Anjili CO 1992. IFN-gamma and delayed-type hypersensitivity
are associated with cutaneous leishmaniasis in vervet monkeys following secondary
rechallenge with Leishmania major. Scand J Immunol (Suppl.) 11:
48-52.
- Pahar B, Li J,
Rourke T, Miller CJ, McChesney MB 2003. Detection of antigen-specific T cell
interferon- expression by ELISPOT and cytokine flow cytometry assays in rhesus
macaques. J Immunol Methods 282: 103-115.
- Palatnik-de-Souza
CB 2008. Vaccines for leishmaniasis in the fore coming 25 years. Vaccine
26: 1709-1724.
- Parrot L, Donatien
A, Lestoquard F 1927. Notes expérimentales sur le bouton d'Orient et
sur la leishmaniose canine viscérale. Arch Inst Pasteur Alger 5:
120-130.
- Pashine A, Valiente
NM, Ulmer JB 2005. Targeting the innate immune response with improved vaccine
adjuvants. Nature Med 11 (Suppl.): 63-68.
- Pitcher CJ, Hagen
SI, Walker JM, Lum R, Mitchell BL, MainoVC, Axthelm MK, Picker LJ 2002. Development
and homeostasis of T cell memory in rhesus macaque. J Immunol 168: 29-43.
- Porrozzi R, Pereira
MS, Teva A, Volpini AC, Pinto MA, Marchevsky RS, Barbosa AA Jr, Grimaldi G Jr
2006. Leishmania infantum-induced primary and challenge infections in
rhesus monkeys (Macaca mulatta): a primate model for visceral leishmaniasis. Trans R Soc Trop Med Hyg 100: 926-937.
- Porrozzi R, Teva
A, Amaral VF, Santos da Costa MV, Grimaldi G Jr 2004. Cross-immunity experiments
between different species or strains of Leishmania in rhesus macaques
(Macaca Mulatta). Am J Trop Med Hyg 71: 297-305.
- Probst RJ, Wellde
BT, Lawyer PG, Stiteler JS, Rowton ED 2001. Rhesus monkey model for Leishmania
major transmitted by Phlebotomus papatasi sandfly bites. Med Vet
Entomol 15: 12-21.
- Pung OJ, Hulsebos
LH, Kuhn RE 1988. Experimental American leishmaniasis and Chagas' disease in
the Brazilian squirrel monkey: cross immunity and electrocardiographic studies
of monkeys infected with Leishmania braziliensis and Trypanosoma cruzi. Internat J Parasitol 18: 1053-1059.
- Pung OJ, Kuhn RE
1987. Experimental leishmaniasis in the Brazilian squirrel monkey (Saimiri
sciureus): lesions, hematology, cellular and humoral immune responses. J
Med Primatol 16: 165-174.
- Ramiro MJ, Zárate
JJ, Hanke T, Rodriguez D, Rodriguez JR, Esteban M, Lucientes J, Castillo JA,
Larraga V 2003. Protection in dogs against visceral leishmaniasis caused by Leishmania infantum is achieved by immunization with a heterologous prime-boost
regime using DNA and vaccinia recombinant vectors expressing LACK. Vaccine
21: 2474-2484.
- Ramos I, Alonso
A, Marcen JM, Peris A, Castillo JA, Colmenares M, Larraga V 2008. Heterologous
prime-boost vaccination with a non-replicative vaccinia recombinant vector expressing
LACK confers protection against canine visceral leishmaniasis with a predominant
Th1-specific immune response. Vaccine 26: 333-344.
- Ranque J, Depieds
R, Nicole RM 1960. Les phénomènes d'immunité dans les leishmanioses. Path Biol 8: 99-107.
- Reed SG, Scott
P 2000. Immunologic mechanisms in Leishmania. MW Cunningham, RS Fujinami
(eds.), Effects of microbes on the immune system. Lippincott Williams
& Wilkins, Philadelphia, p. 537-554.
- Rhee EG, Mendez
S, Shah JA, Wu C, Kirman JR, Turon TN, Davey DF, Davis H, Klinman DM, Coler
RN, Sacks DL, Seder RA 2002. Vaccination with heat-killed Leishmania antigen or recombinant leishmanial protein and CpG oligodeoxynucleotides induces
long-term memory CD4+ and CD8+ T cell responses and protection
against Leishmania major infection. J Exp Med 195: 1565-1573.
- Robinson HL, Amara
RR 2005. T cell vaccines for microbial infections. Nature Med 11 (Suppl.):
25-32.
- Rodríguez-Cortés
A, Ojeda A, López-Fuertes L, Timón M, Altet L, Solano-Gallego
L, Sánchez-Robert E, Francino O, Alberola J 2007. Vaccination with plasmid
DNA encoding KMPII, TRYP, LACK and GP63 does not protect dogs against Leishmania
infantum experimental challenge. Vaccine 25: 7962-7971.
- Saravia NG, Weigle
K, Segura I, Giannini SH, Pacheco R, Labrada LA, Gonçalves A 1990. Recurrent
lesions in humans Leishmania braziliensis infection - reactivation or
reinfection? Lancet 336: 398-402.
- Schneider J, Gilbert
SC, Hannan CM, Degano P, Prieur E, Sheu EG, Plebanski M, Hill AVS 1999. Induction
of CD8+ T cells using heterologous prime-boost immunisation strategies.
Vaccines and vaccination - Part I. Immunol Rev 170: 29-38.
- Sharma SK, Dube
A, Nadeem A, Khan S, Saleem I, Garg R, Mohammad O 2006. Non PC liposome entrapped
promastigote antigens elicit parasite specific CD8+ and CD4+ T-cell immune response
and protect hamsters against visceral leishmaniasis. Vaccine 24: 1800-1810.
- Shearer MH, Dark
RD, Chodosh J, Kennedy RC 1999. Comparison and caracterization of immunoglobulin
G subclasses among primate species. Clin Diagn Lab Immunol 6: 953-958.
- Silveira FT, Lainson
R, Shaw JJ, Garcez LM, Souza AA, Braga RR, Ishikawa EA 1989. Experimental cutaneous
leishmaniasis: I - On the susceptibility of the primate Cebus apella (Cebidae) to the infection caused by Leishmania (Viannia) lainsoni Silveira,
Shaw and Ishikawa, 1987. Rev Soc Bras Med Trop 22: 125-130.
- Silveira FT, Lainson
R, Shaw JJ, Garcez LM, Souza AA, Braga RR, Ishikawa EA 1990. Experimental skin
leishmaniasis: I. Course of the infection in the Cebus apella primate
(Cebidae) caused by Leishmania (V.) braziliensis and L. (L.) amazonensis. Rev Soc Bras Med Trop 23: 5-12.
- Silvestre R, Cordeiro-da-Silva
A, Quaissi A 2008. Live attenuated Leishmania vaccines: a potential strategic
alternative. Arch Immunol Ther Exp 56: 123-126.
- Skeiky YAW, Coler
RN, Brannon M, Stromberg E, Greeson K, Crane RT, Campos-Neto A, Reed SG 2002.
Protective efficacy of a tandemly linked, multi-subunit recombinant leishmanial
vaccine (Leish-111f) formulated in MPL® adjuvant. Vaccine
20: 3292-3303.
- Souza-Lemos C,
de-Campos SN, Teva A, Côrte-Real S, Fonseca EC, Porrozzi R, Grimaldi G
Jr 2008. Dynamics of immune granuloma formation in a Leishmania braziliensis-induced
self-limiting cutaneous infection in the primate Macaca mulatta. J
Pathol 216: 375-386.
- Stäger S,
Smith DF, Kaye PM 2000. Immunization with a recombinant stage-regulated surface
protein from Leishmania donovani induces protection against visceral
leishmaniasis. J Immunol 165: 7064-7071.
- Stober CB, Lange
UG, Roberts MT, Alcami A, Blackwell JM 2005. IL-10 from regulatory T cells determines
vaccine efficacy in murine Leishmania major infection. J Immunol 175:
2517-2524.
- Tabbara KS 2006.
Progress towards a Leishmania vaccine. Saudi Med J 27: 942-950.
- Tapia E, Pérez-Jiménez
E, López-Fuertes L, Gonzalo R, Gherardi MM, Esteban M 2003. The combination
of DNA vectors expressing IL-12 + IL-18 elicits high protective immune response
against cutaneous leishmaniasis after priming with DNA-p36/LACK and the cytokines,
followed by a booster with a vaccinia virus recombinant expressing p36/LACK. Microbes Infect 5: 73-84.
- Teva A, Porrozzi
R, Cupolillo E, Oliveira-Neto MP, Grimaldi G Jr 2005. Responses of Leishmania
(Viannia) braziliensis cutaneous infection to N-methylglucamine antimoniate
in the rhesus monkey (Macaca mulatta) model. J Parasitol 91: 976-978.
- Teva A, Porrozzi
R, Cupolillo E, Pirmes C, Oliveira-Neto MP, Grimaldi G Jr 2003. Leishmania
(Viannia) braziliensis-induced chronic granulomatous cutaneous lesions affecting
the nasal mucosa in the rhesus monkey (Macaca mulatta) model. Parasitology
127: 437-447.
- Titus RG, Bishop
JV, Mejia JS 2006. The immunomodulatory factors of arthropod saliva and the
potential for these factors to serve as vaccine targets to prevent pathogen
transmission. Parasite Immunol 28: 131-141.
- Valenzuela JG,
Belkaid Y, Garfield MK, Mendez S, Kamhawi S, Rowton ED, Sacks DL, Ribeiro JMC
2001. Toward a defined anti-Leishmania vaccine targeting vector antigens:
characterization of a protective salivary protein. J Exp Med 194: 331-342.
- Vanloubbeeck Y,
Jones DE 2004. The immunology of Leishmania infection and the implications
for vaccine development. Ann N Y Acad Sci 1026: 267-272.
- Verthelyi D, Kenney
RT, Seder RA, GamAA, FriedagB, Klinman DM 2002. CpG Oligodeoxynucleotides as
vaccine adjuvants in primates. J Immunol 168: 1659-1663.
- Von Stebut E 2007.
Immunology of cutaneous leishmaniasis: the role of mast cells, phagocytes and
dendritic cells for protective immunity. Eur J Dermatol 17: 115-122.
- Vouldoukis I, Ogunkolade
W, Strazielle L, Ploton I, Monjour L 1986. Susceptibility of Cebus nigrivittatus to Leishmania infantum. J Parasitol 72: 472-473.
- Watkins DI, Kannagi
M, Stone ME, Letvin NL 1988. Major histocompatibility complex class I molecules
of nonhuman primates. Eur J Immunol 18: 1425-1432.
- Webb JR, Campos-Neto,
Ovendale PJ, Martin TI, Stromberg EJ, Badaro R, Reed SG 1998. Human and murine
immune responses to a novel Leishmania major recombinant protein encoded
by members of a multicopy gene family. Infec Immun 66: 3279-3289.
- Webb JR, Kaufmann
D, Campos-Neto, Reed SG 1996. Molecular cloning of a novel protein antigen of Leishmania major that elicits a potent immune response in experimental
murine leishmaniasis. J Immunol 157: 5034-5041.
- Wolf RE 1976. Immune
response to Leishmania tropica in Macaca mulatta. J Parasitol
62: 209-214.
- Zadeh-Vakili A,
Taheri T, Taslimi Y, Doustdari F, Salmanian AH, Rafati S 2004. Immunization
with the hybrid protein vaccine, consisting of Leishmania major cysteine
proteinases Type I (CPB) and Type II (CPA), partially protects against leishmaniasis. Vaccine 22: 1930-1940.
- Zanin FH, Coelho
EA, Tavares CA, Marques-da-Silva EA, Silva Costa MM, Rezende SA, Gazzinelli
RT, Fernandes AP 2007. Evaluation of immune responses and protection induced
by A2 and nucleoside hydrolase (NH) DNA vaccines against Leishmania chagasi and Leishmania amazonensis experimental infections. Microbes Infect
9: 1070-1077.
- Zaph C, Uzonna
J, Beverley SM, Scott P 2004. Central memory T cells mediate long-term
immunity to Leishmania major in the absence of persistent parasites. Nature Med 10: 1104 -1110.
Copyright 2008 - Instituto Oswaldo Cruz - Fiocruz
The following images related to this document are available:
Photo images
[oc08116t3b.jpg]
[oc08116f2.jpg]
[oc08116t2b.jpg]
[oc08116t4b.jpg]
[oc08116t4a.jpg]
[oc08116t3a.jpg]
[oc08116t2a.jpg]
[oc08116f3.jpg]
[oc08116t1.jpg]
[oc08116f1.jpg]
|

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}