 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 103, No. 7, November, 2008, pp. 645-649 ARTICLES Characterization of polymorphisms in the mannose-binding lectin gene promoter among human immunodeficiency virus 1 infected subjects ACR VallinotoI, +; NA MutoI; AEM AlvesI; LFA MachadoI; VN AzevedoI; LLB SouzaII; MOG IshakI; R IshakI IUniversidade
Federal do Pará, Instituto de Ciências Biológicas, Laboratório
de Virologia, R. Augusto Corrêa s/n, 66075-900 Belém, PA, Brasil Financial support: CNPq, PN-DST and Aids/MS Received 22 January
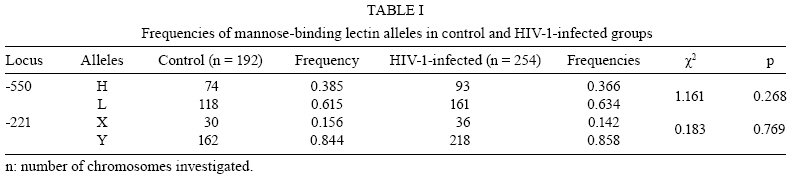
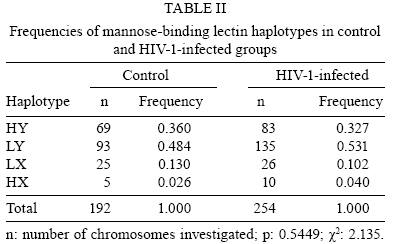
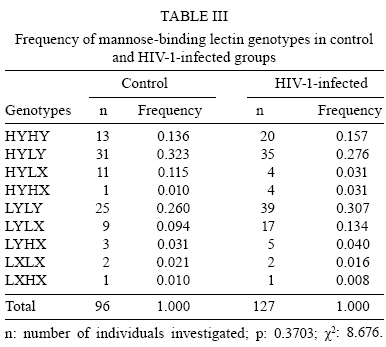
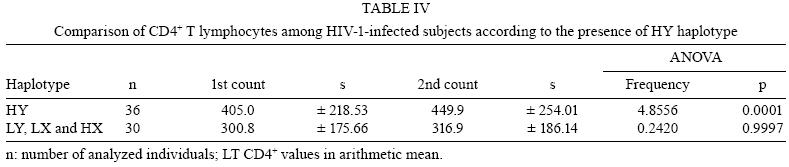
2008 Code Number: oc08117 ABSTRACT The present study investigated the prevalence of mutations in the -550 (H/L) and -221 (X/Y) mannose-binding lectin (MBL) gene promoter regions and their impact on infection by human immunodeficiency virus 1 (HIV-1) in a population of 128 HIV-1 seropositive and 97 seronegative patients. The allele identification was performed through the sequence-specific primer polymerase chain reaction method, using primer sequences specific to each polymorphism. The evolution of the infection was evaluated through CD4+ T-lymphocyte counts and plasma viral load. The allele and haplotype frequencies among HIV-1-infected patients and seronegative healthy control patients did not show significant differences. CD4+ T-lymphocyte counts showed lower levels among seropositive patients carrying haplotypes LY, LX and HX, as compared to those carrying the HY haplotype. Mean plasma viral load was higher among seropositive patients with haplotypes LY, LX and HX than among those carrying the HY haplotype. When promoter and exon 1 mutations were matched, it was possible to identify a significantly higher viral load among HIV-1 infected individuals carrying haplotypes correlated to low serum levels of MBL. The current study shows that haplotypes related to medium and low MBL serum levels might directly influence the evolution of viral progression in patients. Therefore, it is suggested that the identification of haplotypes within the promoter region of the MBL gene among HIV-1 infected persons should be further evaluated as a prognostic tool for AIDS progression. Key words: mannose-binding lectin - genetic polymorphism - human immunodeficiency virus Human immunodeficiency virus (HIV) 1 is one of the most important emerging pathogens. Phylogenetic analysis suggests that HIV-1 appeared between 1915-1941 becoming a public threat 50 years later (Korber et al. 2000). Furthermore, the pathogenesis of HIV-1 infection is complex and influenced by both viral and host factors (Cohen et al. 1997). Mannose-binding lectin (MBL) is a liver-derived, pluripotent serum lectin that plays a role in the human innate immune system, binding with high affinity to mannose or other carbohydrate components present on the surface of viruses, bacteria and yeasts (Kilpatrick 2002). The binding of carbohydrate structures on pathogens may mediate phagocytosis by macrophages, and when this binding is associated with serine proteases it is able to initiate the activation of the complement system through the lectin pathway in an antibody-independent mechanism (Wong et al. 1999, Kilpatrick 2002, Turner 2003). MBL function is directly associated with its serum concentration, which is determined by an interplay between mutations of the promoter and exon 1 (Madsen et al. 1994, 1995). The occurrence of these variants has been associated with MBL serum deficiency and consequently with susceptibility/resistance to infection by pathogens, including HIV-1 (Kilpatrick 2002, Turner 2003). The identification of a mutation in the promoter region showed that serum MBL concentration is also modulated by transcriptional levels (Madsen et al. 1995). Nucleotide substitutions in positions -550 (G-C) and -221 (G-C) provide the variants H(G)/L(C) and Y(G)/X(C), respectively. Several promoter variants that apparently do not alter the levels of MBL were described, but the HY, LY and LX haplotypes are currently associated with high, medium and low levels of circulating MBL in the plasma, respectively (Madsen et al. 1995, Steffensen et al. 2000). Because of linkage disequilibrium among some variants, only seven haplotypes of MBL (HYPA, LYQA, LYPA, LXPA, LYPB, LYQC and HYPD) were identified (Steffensen et al. 2000). The allele dimorphism in the promoter region is known to influence MBL concentration, and the -221 promoter polymorphism is stronger than the -550 one. The present study investigated the relationship of MBL gene polymorphisms in the human host and immunological and viral factors that are capable of influencing the susceptibility to HIV-1 infection and progression to AIDS. SUBJECTS, MATERIALS AND METHODS Population groups examined - Two blood samples were collected five months apart from 127 HIV-1-infected subjects residing in Belém, Pará, Brazil, attending the Reference Unit of Special Infectious and Parasitic Diseases for the determination of CD4+ T- lymphocytes and plasma viral load. All patients were actively receiving antiretroviral therapy (ARVT) at the moment of sample collection, according to the Brazilian consensus of the National Program of STD and AIDS of the Ministry of Health (MS 2004). Ninety percent started therapy within five years and 87% were in their first regimen. The duration of ARVT ranged from 1-7 years. Half of them were in ARVT for no more than five years, with mean and median values of four years. All patients were serum negative for hepatitis C virus (HCV), hepatitis B virus (HBV) and human T-cell lymphotropic virus type 1 (HTLV) infections. Ninety-seven HIV-1 seronegative subjects (screened for antibodies to HIV-1/2, HCV, HBV and HTLV using an enzyme-linked immunosorbent assay) residing in Belém were investigated as controls. All subjects signed a thoroughly-explained consent form at the first visit and all the samples were collected in vacuntainer tubes containing EDTA and directed to the Virus Laboratory of the Universidade Federal do Pará. MBL genotyping - Genomic DNA was extracted from peripheral blood mononuclear cells (PBMC) using the purification kit from Puregene (Gentra Systems, Inc, USA). A sequence-specific primer polymerase chain reaction amplification was performed with a Mastercycle Personal Thermocycler (Perkin-Elmer Cetus Corp., Norwalk CT, USA) to amplify three fragments from the promoter region of the MBL gene [316 pb (H-550)/(L-550), 440 pb (X-221) and 443 pb (Y-221)], following the protocol established previously (Steffensen et al. 2000). The reactions were performed in a final volume of 50 µL containing 500 ng genomic DNA, 225 µM dNTP, 5 pmol primers, 50 mM KCl, 2.5 mM MgCl2, 10 mM Tris-HCl pH 8.3 and 0.5 U of Taq polymerase (Invitrogen, USA). The following primer sequences were used for the genotyping of the promoter region -550 (H/L): (H/L-as) 5' -AACAAATGGGACCGTGCATTGC-3', (H-s) 5' -GCTTACCCAGGAAGCCTGTG-3' and (L-s) 5' -GCTTACCCAGGAAGCCTGTC-3'. Reaction mixtures underwent an initial denaturing step at 95°C for 5 min, followed by 30 cycles of 94°C for 30 s, 59°C for 30 s and 72°C for 45 s, and a final extension period for 5 min. The amplification of the promoter region -221 (X/Y) was performed in a final volume of 50 µL containing 100 ng genomic DNA, 225 µM dNTP, 20 pmol primers, 50 mM KCl, 2.5 mM MgCl2 , 10 mM Tris-HCl pH 8.3 and 0.5 U of Taq polymerase (Invitrogen, USA). Primer sequences were: (Seq-s) 5' -CCTGCCAGAAAGTAGAGAGG-3', (Y-as) 5' -CTGGAAGACTATAAACATGCTTTCC-3' and (X-as) 5' -GGAAGACTATAAACATGTTTCG-3'. Reaction mixtures underwent an initial denaturing step at 95°C for 5 min, followed by 35 cycles of 95°C for 30 s, 59°C for 45 s and 72°C for 45 s, and a final extension period for 5 min. The alleles H, L, Y and X were additionally amplified with primers Y-as (-221) and L-s (-550) or Y-as (-221) and H-s (-550) to determine the cis/trans structural configuration of the haplotype, as previously described (Steffensen et al. 2000). Quantification of plasma viral load and determination of CD4+ T-lymphocytes - The plasma viral load was determined via the Nasba method using the NucliSens Reader equipment and the NucliSensTM kit and followed the directions of the manufacturer (Nasba Diagnostics, Organon Teknika, Boxtel, Netherlands). Whole blood samples were processed within 4 h of collection for the determination of the T-lymphocytes subset counts by flow cytometry (FacsCount, Becton & Dickinson, USA) using the FacsCountTM Reagents immunomonitoring kit and following the protocol recommended by the manufacturer (Becton Dickinson, USA). Statistical analysis - The allelic, haplotype and genotype frequencies in both patient and control groups were estimated by directed counting. The comparison of all-elic, haplotype and genotype frequencies between the patient and control groups was performed using the 2 test by the Tool for Population Genetic Analyses software 1.3v (Miller 1999). The association analysis between the genetic polymorphism of the promoter region of MBL gene, plasma viral load and CD4+ T-cell count were performed using the Student's t-test and ANOVA using the BioEstat software, version 4.0 (Ayres et al. 2005). RESULTS The allele L (-550 MBL gene region) was detected with a high prevalence in both seronegative and HIV-1-infected subjects [0.615 and 0.634, respectively (Table I)]. The allele Y (-221 MBL gene region) was the most frequent in both groups (0.844 and 0.858, respectively). The differences in allelic frequencies (L versus H and Y versus X) were not statistically significant (p = 0.268 and p = 0.769, respectively). Table II describes the frequencies of MBL haplotypes. The LY haplotypes was the most prevalent (0.484 and 0.531, respectively) and the HX haplotype was least prevalent (0.026 and 0.040, respectively), but the differences among the frequencies between the HIV-1-infected and control groups were not statistically significant. Nine genotypes were described and present in both groups considered (Table III). Their frequencies ranged from 0.010-0.323 in the control group and from 0.008-0.307 in the HIV-1-infected patients. The differences in the genotype frequencies were not statistically significant. The mean values of both CD4+ T lymphocyte counts (collected 5 months apart) obtained from subjects carrying the HY haplotype in homozygous or heterozygous forms were significantly different compared to those carrying the LY, LX and HX haplotypes. In addition, HIV-1-infected subjects presented a significant increase of CD4+ T lymphocytes from the first to the second quantification (Table IV). The presence of either the homo or heterozygous genotype HY (HY/HY or HY/*) apparently conferred higher counts of CD4+ T lymphocytes that were statistically significant when compared to other genotypes (Table V). Measurements of plasma viral load were distributed according to subjects with HY haplotypes and compared to subjects carrying LY, LX and HX haplotypes. HIV-1-infected patients who carried haplotype HY showed a significant reduction (p = 0.0062) in the plasma viral load, which was not observed among HIV-1-infected patients carrying other haplotypes (Table VI). Additionally, when promoter and exon 1 mutations were matched, we observed that the 64 patients carrying the halotypes LYA/O, LYO/O, LYO/LXA, HY/LXO, LY/LXO (correlated with low serum levels of MBL) had a significantly higher viral load (52.7 copies/mm3; log10 = 4.72) compared to the 63 patients carrying the haplotypes HYA/A, HY/LYA, LYA/A, LY/LXA, HYO/O, HYA/LYO, HYA/LXO (correlated to medium and high levels of MBL) (21.8 copies/mm3; log10 = 4.33; p < 0.05). DISCUSSION The present study described the distribution of allelic variants of the promoter region of the MBL gene and compared this distribution among HIV-1-infected and uninfected subjects. MBL is an important serum protein involved in the innate immune response, which is able to trigger complement activation (Wong et al. 1999, Kilpatrick 2002, Turner 2003). The presence of mutations in exon 1 of the MBL gene has been associated with the occurrence of immunodeficiency and chronic infection diseases (Kilpatrick 2002, Turner 2003). In our study, the variant MBL*B was clearly associated with higher plasma viral load levels (Vallinoto et al. 2006). The frequency distribution of allelic variants of the promoter region of the MBL gene described in this study was similar to the frequencies reported by Steffensen et al. (2000) and Michington et al. (2002) among Danish and Australian blood donors, respectively, as well as by Hakozaki et al. (2002) among HBV carriers and uninfected subjects in Japan. The similarity of the frequencies may be a usual convergence of the haplotypes among different population groups even when they are from distinct geographic locations. A small difference was observed in the frequency in position -550 of the promoter region of the MBL gene. This suggests a possible predisposition to HIV-1 infection due to the high frequency of HIV-infected subjects carrying the allele L in homozygous form, but no significant differences in the frequencies of alleles, haplotypes or genotypes among HIV-1-infected and uninfected subjects was reported. Nielsen et al. (1995) and Pastinen et al. (1998) have shown that HIV-1-infected subjects presented lower serum levels of MBL compared to uninfected subjects and, according to Mangano et al. (2008), MBL plasma concentration is associated with the rate of AIDS progression when considering vertical transmission of the virus. This model was not supported by Senaldi et al. (1999). No significant correlation was observed between the level of serum MBL and the time of detection of antibodies to HIV-1, the development of AIDS, the diagnosis of AIDS or death (Nielsen et al. 1995, Garred et al. 1997a, b, Prohaszka et al. 1997, Mcbrige et al. 1998). Low levels of serum MBL have been associated primarily with a significant increase (Prohaszka et al. 1997) or reduction (Senaldi et al. 1999) in survival time after the diagnosis of AIDS, as well as with the increase of the time between seroconversion and development of AIDS and death (Maas et al. 1998, Mangano et al. 2008). Serum levels of the MBL protein have a direct association with the polymorphisms of the promoter region and with mutations in exon 1 of MBL gene. The absence of statistical association of the frequencies of alleles, haplotypes and genotypes in the present study suggests that the mutations in the promoter region, which are related to medium and low levels of MBL serum protein, are not clearly associated to the susceptibility/resistence to infection by HIV-1, rate of infection or progression to AIDS. It has been suggested that the deficiency of MBL protein increases the susceptibility to HIV-1 infection and the progression of disease (Nielsen et al. 1995, Garred et al. 1997a, b, Prohaszka et al. 1997, Mcbrige et al. 1998, Pastinen et al. 1998). In Belém, a significant increase of the MBL*B variant among HIV-1-infected subjects was observed (Vallinoto et al. 2006). Although the genotype B/B was six times more frequent, neither allele nor genotype differences were significant. MBL gene polymorphisms may have a direct role in HIV-1 infection in which low serum levels of MBL protein contribute to the increase of plasma viral load (Garred et al. 1997a, b). In the present study, infected subjects carrying the HY haplotype clearly reduced their plasma viral load between the two laboratory counts performed three to six months apart, while patients carrying haplotypes LX and LY did not. These results are similar to what has been described previously in Belém, in which the polymorphism in codon 54 of the MBL gene leads to a higher plasma viral load among HIV-1-infected subjects (Vallinoto et al. 2006). Additionally, when matching the polymorphism results of the promoter and exon 1 regions, we identified a significant association between high viral load and the presence of genotypes correlated to low MBL serum levels, supporting our previous report showing the direct influence of the structural mutation on exon 1 regarding HIV-1 viral load (Vallinoto et al. 2006). Thus, the results obtained suggest that the low serum levels of MBL could contribute to an inefficient elimination of the virus and a consequent increase of viral load. Nielsen et al. (1995) and Pastinen et al. (1998) described an absence of correlation between the serum concentration of MBL and the decline of CD4+ T lymphocytes among HIV-1 infected subjects. The present results describe a significant increase of CD4+ T lymphocytes among HIV-1-infected subjects carrying haplotype HY either in homo or heterozygous form. It is possible that the reduction of plasma viral load decreases the level of T lymphocyte infection. Higher concentrations of circulating MBL favor the activation of the complement system and indirect blocking of viral replication. Michington et al. (2002) confirm that the presence of the HY and LY haplotypes are associated to higher concentrations of MBL in contrast to the low circulating levels associated with the LX haplotype. The role of MBL in HIV-1-infected subjects is still a controversial matter. It is possible that the identification of the polymorphism in the promoter region could be used as a biomarker for the monitoring of antiretroviral therapy in conjunction with CD4+ T lymphocyte counts and plasma viral load. Furthermore, there is a need to evaluate the actual impact of MBL serum deficiency on the course of HIV-1 infection, particularly in light of the potential use of recombinant MBL as a therapeutic tool. ACKNOWLEDGMENTS To all of the subjects involved in this study. REFERENCES
Copyright 2008 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc08117t1.jpg] [oc08117t2.jpg] [oc08117t3.jpg] [oc08117t6.jpg] [oc08117t5.jpg] [oc08117t4.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}