 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 103, No. 7, November, 2008, pp. 678-682 ARTICLES Diversity of Chromobacterium violaceum isolates from aquatic environments of state of Pará, Brazilian Amazon LT Dall'AgnolI; RN MartinsII; ACR VallinotoII; KTS RibeiroI, + ILaboratório
de Microbiologia e Imunologia Financial support: CNPq, Programa de Pós-Graduação Biologia de Agentes Infecciosos e Parasitários/UFPA Received 16 May
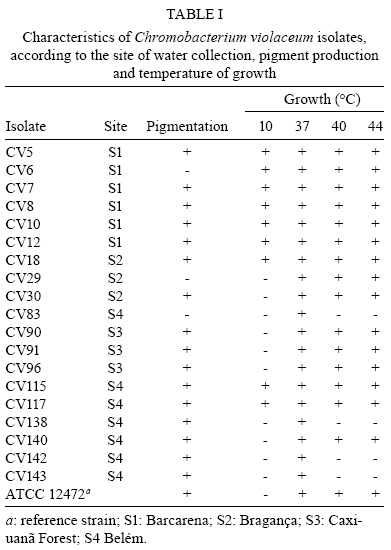
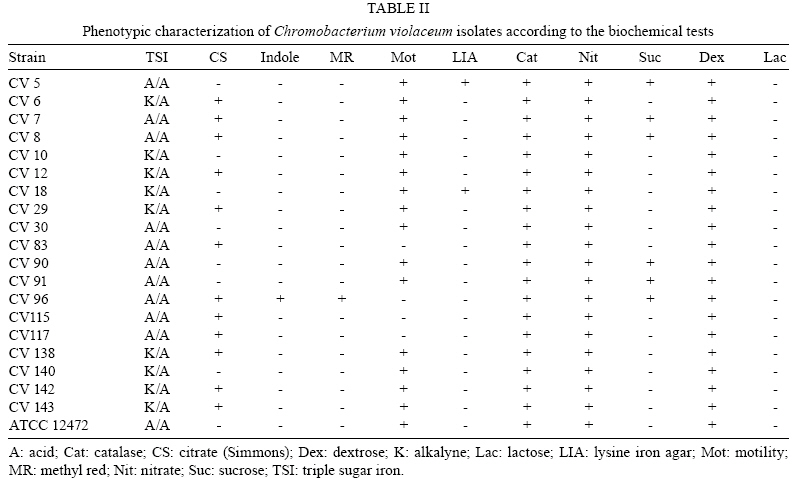
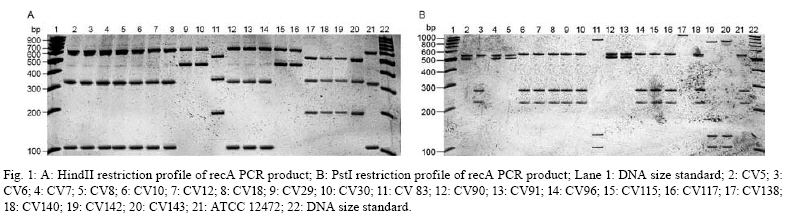
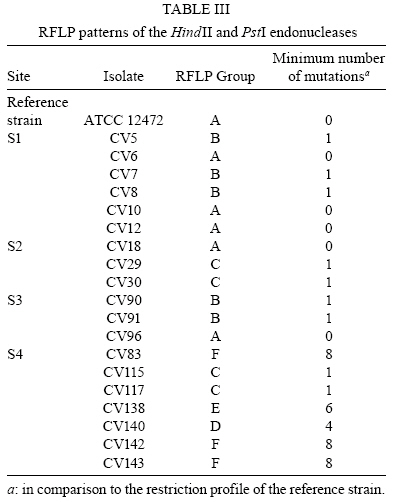
2008 Code Number: oc08123 ABSTRACT The present study intended to characterize the phenotypic and genetic diversity of Brazilian isolates of Chromobacterium violaceum from aquatic environments within the Amazon region. Nineteen isolates showed morphological properties of C. violaceum and the majority grew at 44°C. Low temperatures, in contrast, showed to be inhibitory to their growth, as eleven isolates did not grow at 10ºC and nine did not produce pigmentation, clearly indicating an inhibition of their metabolism. The largest variation among isolates was observed in the citrate test (Simmons), in which 12 isolates were positive, and in the oxidation/fermentation of sucrose, with six positives isolates. Chloramphenicol, gentamicin and sulfonamides efficiently inhibited bacterial growth. Amplified products of the recA gene were digested with HindII or PstI, which produced three or four restriction fragments patterns, respectively. The combined analysis arranged the isolates into six genospecies. The higher diversity observed in Belém (genotypes C, D, E and F) may be a consequence of intense human occupation, pollution of the aquatic environment or due to the higher diversity of the environments sampled in that region. In conclusion, a high level of genetic and phenotypic diversity was observed, and four new genospecies were described. Key words: Chromobacterium violaceum - Recombinase A - RFLP - diversity - Amazon region Chromobacterium violaceum was first described by Bergonzini in 1880 (Sneath 1984) and belongs to the family Neisseriacea of -Proteobacteria. It is a Gram-negative, heterotrophic, flagellated, free-living organism which lives in a variety of ecosystems in tropical and subtropical regions, including the soil and water of the Amazon region (Hungria et al. 2004). The main feature of this microorganism is the production of a purple pigment named violacein (Antônio & Creczynski-Pasa 2004, Dessaux et al. 2004), although non-pigmented strains have also been reported (Sivendra & Tan 1977). Violacein possesses anti-leishimanial (Leon et al. 2001), anti-viral (Andrighetti-Fröhmer et al. 2003), anti-tumoral (Ueda et al. 1994, Melo et al. 2000) and anti-Mycobacterium tuberculosis (de Souza et al. 1999) activities. Other properties of C. violaceum include the production of cyanide (Michaels & Corpe 1965), the solubilization of gold (Faramarzi et al. 2004), the production of chiti-nolytic enzymes (Chernin et al. 1998), the synthesis of bioplastics (Steinbüchel et al. 1993) and environmental detoxification (Carepo et al. 2004). Although it is a valuable biotechnological resource, C. violaceum is a highly virulent opportunistic pathogen to humans and animals (Chen et al. 2003, Brito et al. 2004, Dias et al. 2005, de Siqueira et al. 2005). The genome of C. violaceum, strain ATCC 12472, was sequenced by the Brazilian National Genome Project Consortium (de Vasconcelos et al. 2003), but few studies have investigated the diversity of indigenous isolates in Brazil or in the Amazon region (Hungria et al. 2005, Lima-Bittencourt et al. 2007). Recombinase A (RecA) is a multifunctional protein involved in general recombination, DNA repair and the SOS response, and it is highly conserved among eubacteria (Cox 2003). Several studies have shown that recA can be used as a molecular tool to study diversity within the Erwinia genus (Waleron et al. 2002), C. violaceum (Scholz et al. 2005), Ochrobactrum anthropi (Scholz et al. 2006) and the Burkholderia cepacia complex (Seo & Tsuchiya 2004), despite this gene's high degree of nucleotide diversity (Casati et al. 2004). The present study aims to further characterize the phenotypic and genetic diversity of Brazilian isolates of C. violaceum from aquatic environments within the Amazon region. MATERIALS AND METHODS Isolates from natural water resources and reference strain - Water samples were collected from domestic wells (less than 15 m in depth) in Barcarena (site 1) and in Vila Bonifácio, municipality of Bragança (site 2). Samples were also collected from rivers within the National Forest of Caxiuana (site 3) and in Belém, the capital of Pará (site 4). Nineteen isolates of C. violaceum were recovered from superficial and underground water using the Membrane Filter (MF) and Most Probable Number (MPN) technique, according to the Standard Methods for the Examination of Water and Wastewater (APHA 2005). The MF technique was performed by the filtration of water samples through a sterile cellulose ester membrane with 0.45 µm pores. Membranes were transferred to m-Endo-type media with incubation at 37ºC for 24 h. The MPN technique consisted of the inoculation of sequential volumes of water samples in Lactose Broth, followed by incubation at 37ºC for 24-48 h. The colonies that presented a violet pigmentation in the isolation medium were transferred to Agar Stock (storage medium). Prior to biochemical or molecular testing, isolates were grown in Nutrient Broth (NB) for 24 h at 37ºC and placed in Nutrient Agar to re-isolate and confirm the anoxic condition of the culture. The reference strain ATCC 12472 was obtained from the Brazilian Collection of Industrial and Environmental Microorganism. Phenotypic characterization - Isolates were transferred to NB and incubated at 10, 37, 40 and 44°C for 24-48 h. Cultures positive at 37°C were inoculated to test growth ability and pigment production. Each isolate was also inoculated in triple sugar iron agar and from this inoculated to indole, methyl red, citrate (Simmons), lysine iron agar, catalase, motility, nitrate and oxidation/fermentation of sucrose, dextrose and lactose. Susceptibility to antimicrobials - The susceptibility test was performed in Mueller-Hinton agar using the Kirby-Bauer disc diffusion method, according to Bauer et al. (1966). The antimicrobials tested were cefoxitin (30 µg), chloramphenicol (30 µg), gentamicin (10 µg), penicillin (10 U) and sulfonamides (300 µg). The results were determined by measuring the inhibition of the growth halo and comparing with the standard values established by the Clinical and Laboratory Standard Institute Manual (NCCLS 2003). DNA extraction - Genomic DNA was extracted from bacterial culture after incubation for 48 h in NB at 35ºC using the SDS/proteinase K based method described by Ausubel et al. (2003), with the addition of 5% RNAse for 1 h at 55ºC (within the proteinase K). RecA amplification PCR - The primer pair recA-viol-f' (5' -AAGACAAGAGCAAGGCGCTGGC-3') and recA-viol-r (5' -TCGAAGGCGTCGTCGGCGAAC-3') (Scholz et al. 2005) generated a 1040 bp fragment. PCR was performed in an Eppendorf Mastercycler Personal thermocycler in a 50 µL mix with 10 mol of each primer and a program of an initial cycle of denaturation at 94°C for 3 min; 30 cycles of denaturation at 94°C for 30 s, annealing at 67°C for 30 s and elongation at 72°C for 90 s; a final extension cycle at 72°C and a final soak at 4°C for 5 min completed the run. Each PCR product (10 µL) were visualized in a 0.8% (in TAE buffer) agarose gel eletrophoresis after staining with ethidium bromide. RecA RFLP analysis - Each PCR product was digested in separate reactions with the two restriction endonucleases PstI (Invitrogen, USA) and HindII (Sigma, USA). Digestion was performed in a mix of 50 µL with 5 U of each enzyme and 15 µL of each PCR product for 2 h, using the conditions recommended by the manufacturer. Restriction fragments were separated in a 10% polyacrylamide gel electrophoresis and documented by photography following silver and sodium hydroxide staining. Dendrogram construction - The enzymatic digestion results were analyzed with the Network 4.5 software (http://www.fluxus-engineering.com/sharepub.htm#a1), using the median joining algorithm (Bandelt et al. 1999) to construct a dendrogram of the relationship between isolates and the reference strain. RESULTS Phenotype characterization - Nineteen isolates and the reference strain were characterized according to their pigment production at 37ºC and their ability to grow at temperatures ranging from 10-44ºC (Table I). All the isolates showed morphological properties of C. violaceum (Sneath 1984). Strains CV6, CV29 and CV83 lost the ability to produce pigmentation during storage and CV83, CV96, CV115 and CV117 were motility negative. Biochemical identification was confirmed by the amplification of the fragment of recA gene. All C. violaceum strains studied displayed similar properties in our biochemical characterization (Table II), with the exception of strains CV96 (positive in the indole and methyl red tests), CV5 and CV18 (positive in the lysine test). The largest variation was observed in the citrate test (Simmons), in which 12 isolates were positive, and in the oxidation/fermentation of sucrose, with six positives isolates. Susceptibility to antimicrobials - Chloramphenicol, gentamicin and sulfonamides were sufficient to inhibit the growth of both the isolates and the reference strain. CV140 was only partially inhibited by gentamicin and CV83 was the sole isolate inhibited by cefoxitin. All isolates were resistant to penicillin. RecA amplification and RFLP analysis - The amplification of recA generated a fragment of 1,040 bp (from the recA gene of 1,057 bp). Amplified products were digested with HindII or PstI, which produced three or four restriction fragments patterns, respectively. The isolates were arranged into six genospecies (Figs 1, 2, Table III). Isolates with no mutation in any restriction site (RS) of the two enzymes were classified into group A. Those isolates with at least one mutation in the PstI RS but none in HindII were placed into group B. Alternatively, those that had at least one mutation in the HindII RS and none in PstI were placed into group C. The single isolate with no mutation in the PstI RS but four mutations in HindII was placed in group D. Group E was composed by the isolate that had at least four mutations in RS of HindII and at least two in PstI. Finally, group F consisted of those isolates that had at least four mutations in the RS of both HindII and PstI. DISCUSSION C. violaceum is a saprophitic organism that usually grows at a maximum temperature of 40ºC (Hungria et al. 2005); however, the majority of indigenous isolates are able to grow at 44ºC, which was seen with the isolates examined in this study. This result is probably related to the high temperatures of the region from which these organisms were isolated (Hungria et al. 2005). One particularly interesting observation from this study was that four isolates from site 4 exhibited reduced growth rates in temperatures above 37°C. Such a growth defect could be a consequence of the longer duration of storage of those isolates. Although the minimum temperature allowing growth of the species is usually between 10-15ºC (Sneath 1984), other isolates of the region (Hungria et al. 2005) showed no growth under 15ºC. On the other hand, 11 isolates exhibited growth after incubation at 10ºC and none of them produced pigment under these conditions. It is possible that low temperatures are more restrictive to C. violaceum and that the production of violacein is not essential for the survival of the bacteria (Efthimion & Corpe 1969). A larger phenotypic variation was observed under the citrate test and the oxidation/fermentation of sucrose, which reflects the diversity of metabolic pathways to obtain nutrients from the environment. The biochemical results indicate the great environmental adaptability and tolerance to stress of C. violaceum (Creczynski-Pasa & Antônio 2004, Hungria et al. 2004). Human infections of C. violaceum are uncommon and of different clinical and laboratory diagnosis (Scholz et al. 2005). The typical infection occurs as a short-duration, highly virulent disease; therefore, the need for a rapid diagnosis and susceptibility profile to antimicrobials is urgent (Dias et al. 2005). When compared to in vitro data, the results obtained with the antimicrobials sensitivity test are very similar to those observed in other studies (Lee et al. 1999). In silico studies of the genome (Fantinatti-Garboggini et al. 2004) detected a probable gene of chloramphenicol resistance; however, resistance to this antibiotic was not observed in vitro. The primers used in the present study were sufficient to biochemically identify the recA gene through PCR amplification of a 1,040 bp fragment. RFLP analysis was previously described as a useful genetic marker to indicate bacterial variability both within and among species (Waleron et al. 2002, Casati et al. 2004, Seo & Tsuchiya 2004, Scholz et al. 2006). Genotype A was described in most of the sites sampled (1, 2 and 3). Site 3 is a National Florest reserve, which is supposedly not influenced by the presence of humans. The higher diversity observed in Belém (genotypes C, D, E and F) may be a consequence of intense human occupation, pollution of the aquatic environment or possibly a reflection of the higher diversity of environments sampled. A similar degree of diversity was observed in isolates from the state of Amazonas (Hungria et al. 2005) which suggests that high diversity is an intrinsic characteristic of the species found within the Amazon region. The present study showed one new restriction profile for the HindII enzyme and three for PstI. Previously described strains with one restriction profile in HindII and two in PstI were absent (Scholz et al. 2005). These results demonstrate the great variability, both phenotypic and genetic, of the species in the Amazonian aquatic environment, and therefore they are important for a better understanding of C. violaceum adaptability. Furthermore, this report may form a basis for further studies exploring all the potentialities that this bacterium offers. ACKNOWLEDGMENTS To the Laboratório de Polimorfismo de DNA/UFPA, for material support, especially to Rafael Silva Rocha and Ricardo Souza, and to Professor Ricardo Ishak, for reviewing the manuscript. REFERENCES
Copyright 2008 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc08123f1.jpg] [oc08123t1.jpg] [oc08123t2.jpg] [oc08123f2.jpg] [oc08123t3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}