 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 103, No. 7, November, 2008, pp. 683-689 ARTICLES The redox potential interferes with the expression of laminin binding molecules in Bacteroides fragilis Eliane de Oliveira FerreiraI, +; Edwin Alexander YatesII; Morris GoldnerIII; Rossiane Cláudia VommaroIV; Fernando Costa e Silva FilhoIV; Débora Barreiros PetrópolisIV; Regina MC Pilotto DominguesI ILaboratório
de Biologia de Anaeróbios, Departamento de Microbiologia Médica,
IMPPG Financial support: CAPES, FAPERJ, CNPq, PRONEX Received 1 June
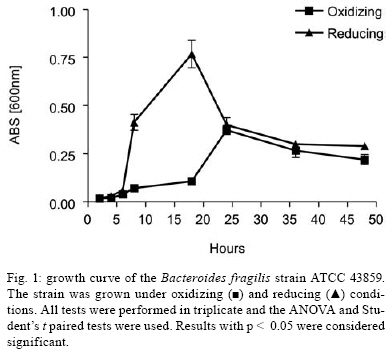
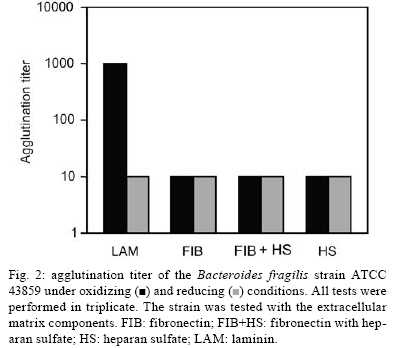
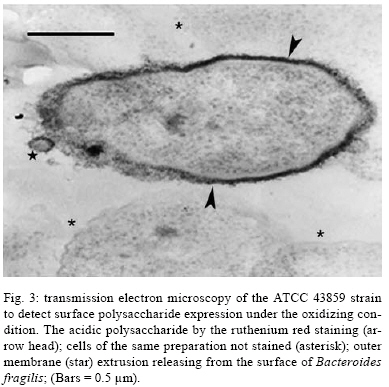
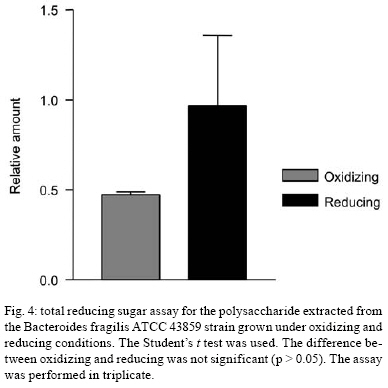
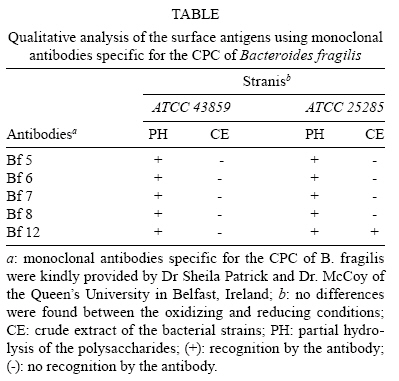
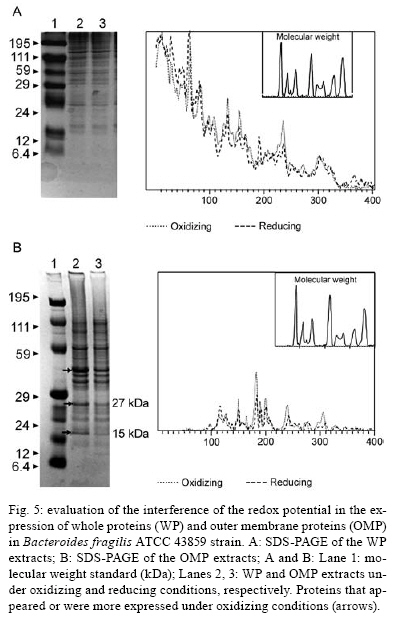
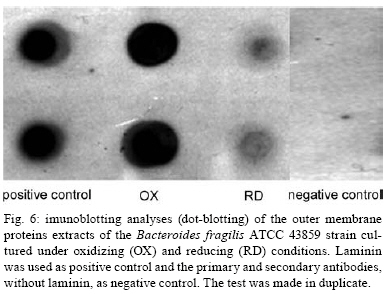
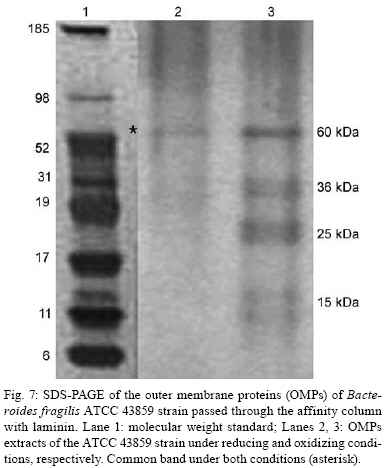
2008 Code Number: oc08124 ABSTRACT The Bacteroides fragilis ATCC strain was grown in a synthetic media with contrasting redox potential (Eh) levels [reduced (-60 mV) or oxidised (+100mV)] and their adhesion capacity to extracellular matrix components was evaluated. The strain was capable of adhering to laminin, fibronectin, fibronectin + heparan sulphate and heparan sulphate. A stronger adherence to laminin after growing the strain under oxidising conditions was verified. Electron microscopy using ruthenium red showed a heterogeneous population under this condition. Dot-blotting analyses confirmed stronger laminin recognition by outer membrane proteins of cells cultured at a higher Eh. Using a laminin affinity column, several putative laminin binding proteins obtained from the cultures kept under oxidising (60 kDa, 36 kDa, 25 kDa and 15 kDa) and reducing (60 kDa) conditions could be detected. Our results show that the expression of B. fragilis surface components that recognise laminin are influenced by Eh variations. Key words: Bacteroides fragilis - adhesion - redox potential - laminin binding proteins - outer membrane proteins Bacteroides fragilis is a Gram-negative obligate anaerobic bacterium and a common member of the human microbiota (Jousiemies-Somier 2002), but it is also an important opportunistic pathogen. This bacterium is the most frequently isolated anaerobic species from human intra-abdominal infections, and this represents a serious clinical problem, especially when abscesses are formed (Finegold 1995). Although several factors have been described, the pathogenicity of this microorganism is still not completely understood. The capsular polysaccharide complex (CPC) is a crucial factor in the virulence potential of the species (Comstock et al. 1999, Coyne et al. 2000, 2001). B. fragilis modulates its surface antigenicity, generating distinct CPC combinations (Krinos et al. 2001). High variation in the polysaccharide expression of B. fragilis has already been demonstrated using monoclonal antibodies (Patrick et al. 1999). Recent analyses of the genome sequences revealed DNA inversion regions. These results suggested that the ability of B. fragilis to colonise several host environments, behaving either as a member of the microbiota or a pathogen, might be linked to switching of polysaccharide biosynthesis (Krinos et al. 2001). The extracellular matrix (ECM) is a stable macromolecular structure underlying epithelial and endothelial cells and surrounding connective tissue cells. In the ECM, collagens, proteoglycans and structural glycoproteins, such as fibronectin and laminin, are found and these molecules are usually exploited for colonisation by microbes (Ljungh et al. 1996). The cell surface adhesins that mediate such recognition are termed "microbial surface components recognising adhesive matrix molecules" (MSCRAMMS). In the past, several microorganisms have been shown to express these molecules (Ljungh at al. 1996, Lähtennmaki et al. 1998, Crago & Koronakis 1999). Many bacterial virulence factors are well regulated with their expression linked to several environmental signals, such as temperature, pH, osmolarity, carbon source and iron levels (Sijbrandi et al. 2005). To the obligate anaerobic bacteria, the redox potential (Eh) represents an important environmental parameter to their survival. It has been shown that, when grown at different Eh levels, B. fragilis can change its state from commensal to pathogenic, becoming invasive to HeLa cells (Goldner et al. 1993). In the present investigation, we searched for B. fragilis properties regulated by Eh. We investigated interference with adhesion to ECM components and with the expression of proteins and surface polysaccharides. Our results showed that the adhesion to laminin is influenced by Eh, and we report that several putative binding proteins were more highly expressed under oxidising conditions by ATCC 43859 strain. MATERIAL AND METHODS Bacterial strain - A reference strain, ATCC 43859, was selected for this study. ATCC strain 25285 was used as a control in the assays with monoclonal antibodies. Both strains were obtained from the American Type Culture Collection, Rockville, MD, USA. Culture conditions - The strain was first grown in brain heart infusion (BHI, Sigma Co), consisting of previously reduced and anaerobically sterilised (PRAS) medium (37ºC, 18 h) and blood agar supplemented with hemin (5 mg/mL; Sigma Co) and vitamin K (0.1 mg/mL; Sigma Co), under anaerobic conditions (80% N2; 10% CO2; 10% H2) at 37ºC (Gumbiner 1996). After 48 h of incubation, colonies were selected and grown in BHI-PRAS at 37ºC for 18 h. After growing in BHI-PRAS medium (37ºC, 18 h), 300 µL of the culture (109 CFU/mL) was transferred to the oxidising (Eh7 + 100 mV) and reducing (Eh7 - 60 mV) medium described by Goldner et al. (1993). The tubes were incubated for 18 h at 37ºC. The bacterial suspension (109 CFU/mL) was prepared after centrifugation (3000 g) and washed twice with 0.1 M PBS (pH 7.2). Bacterial growth - A growth curve was constructed under both conditions. One milliliter of the bacterial culture was taken at 2, 4, 6, 8, 18, 24 and 48 h, and the absorbance at 660 nm (Pharmacia Biothec, Ultrospec 2000) was measured. All experiments were performed in triplicate. For statistical analysis, the ANOVA and the Student's t paired tests, both from SPSS, Version 1.3 (p < 0.05), were used. Latex beads - Latex beads (Sigma, diameter: 0.82 µm) were prepared according to Naidu et al. (1988). Briefly, a latex suspension was mixed with 3 mL of a 0.17 M glycine-NaOH buffer (pH 8.2), centrifuged at 3000 g and washed twice with the same buffer. The pellet was mixed with 3 mL of the glycine buffer and ECM components: laminin [17 µg/mL, Sigma Co, Tumor Engelbreth-Holm-Swarm (EHS)], fibronectin (33 µg/mL, Sigma Co), fibronectin (33 µg/mL) + heparan sulphate (33 µg/mL, Celsus Laboratories, Inc) and heparan sulphate (33 µg/mL). The suspension was shaken horizontally (80 rpm, 30ºC, 14 h), centrifuged, and washed with 2 mL of glycine buffer (0.01% BSA). The latex beads were kept at 4ºC. Agglutination assays - The assays were performed according to Nagy et al. (1994). For the test, 10 µL of the bacterial suspension, started from 109 CFU/mL and following a 10-4 fold dilution series, and 10 µL of the corresponding coated latex suspension were dropped onto a glass slide and mixed gently. The reaction was read after 2 min. As a negative control, the bacteria were mixed with the glycine-NaOH buffer (pH 8.2) or with BSA (2 mg/mL in the same buffer) latex beads. The titer was defined as the last dilution to cause agglutination. The strain was tested in triplicate. Surface polysaccharide expression - Transmission Electron Microscopy (TEM) - For detecting acidic surface polysaccharide expression, ruthenium red staining was used. The cells were grown as described before, washed with PBS (0.1 M, pH 7.2) and the pellet resuspended in equal volumes of 2.5% (v/v) glutaraldehyde, RR solution (10 mg/mL) and 0.1 M cacodylate buffer (pH 7.2) for 1 h at rt. Cells were fixed with 4% osmium tetroxide (OsO4), RR solution (1%) and 0.1 M cacodylate buffer for 1 h at rt. The suspensions were washed (3 x), dehydrated in acetone (30-100%) and then embedded in Epon/acetone (1:1) for 24 h at rt and in Epon (2 days/60ºC). Ultrathin sections were made using an ultramicrotome (Reichert Ultracuts-Leica), and micrographs were taken on a Ziess 900 electron microscope. Total polysaccharide dosage - After growing the strain under the previously described culture conditions, 5 mL was centrifuged (4000 x g) and washed (6 x) to remove all sugar derived from the medium; the bacteria were sonicated (2 min, 4ºC) and then washed (4000 g; 2 x) to remove any sugar present in the cytosol. The samples were freeze-dried, then hydrolysed (2 M HCl, 100ºC, 3 h) and freeze-dried again. A sugar assay, using neocuproine, was performed according to the method described by Chaplin and Kennedy (1994). The Student's t test was used for comparing results from oxidising and reducing conditions. Detection of surface antigens by monoclonal antibodies - To evaluate the polysaccharide antigens exposed on the bacterial surface, ATCC strain 43859 and ATCC strain 25285 were used. The crude total polysaccharide extract obtained was hydrolysed in a 50% acetic acid solution at 100ºC for 30 min and freeze-dried prior to probing using an ELISA assay. For the assay, described by Patrick et al. (1999), the freeze-dried hydrolysed polysaccharide extract was dissolved and 100 µL was applied to each of 96 wells and, after 24 h at 4ºC, each well was blocked with 2% BSA (Sigma Co) in 0.1 M PBS pH 7.2 for 1 h at 4ºC. Fifty microlitres of each monoclonal antibody was added and the plate was incubated for another hour at 4ºC. After washing, the peroxidase-conjugated secondary antibody (1:1000 in 0.1 M PBS pH 7.2 with 2% BSA) was added and incubated for 1 h at 4ºC. To develop the assay, ortho-phenyl-diamine (Sigma Co) in 5 mL 0.1 M PBS with 10 µL of sodium peroxide was used. Assay development was stopped with 0.05 M sulphuric acid in 0.5 M PBS. The plate was read in a spectrophotometer (BIORAD model 3550) at 450 nm. Whole proteins (WP) - After cultivation of the strain, 1 mL of the culture was centrifuged (4000 g), washed twice with PBS and the pellet was mixed with 100 µL of 0.25 M Tris-HCl, 0.192 M glycine, 0.1% SDS, pH 8.5, and stored at -20ºC (Taylor et al. 1986). SDS-PAGE analysis was carried out in a Bis-tris Gel (4-12% Nu PageTM, 1 mm x 12 wells) in a vertical slab gel apparatus (20 mA, 100V) (Laemmili 1970). A molecular weight standard of proteins (Invitrogen) was used. All gels were silver stained. Densitometry analysis (Molecular Analyst version 1.6, Bio-Rad) was performed. Outer membrane proteins (OMP) - After cultivation of the strain, 500 mL of the culture was centrifuged (4000 g), washed (0.1 M PBS; 2 x) and resuspended in 10 mM Tris-HCl, pH 8.0, 1 mM EDTA and 1mM β-Mercaptoethanol (Sigma Chem. Co). The cells were sonicated (2 min, 4ºC) and the intact cells were removed by centrifugation (4000 g) for 10 min. The supernatant was centrifuged (100.000 g, 30 min, 4ºC) and the sediment was treated twice with a 0.3% Sarcosyl (Sigma Chem. Co) solution, centrifuged (7500 g, 4ºC), and then resuspended in 0.01 M phosphate buffer (Bölin et al. 1982). The proteins were kept at -20ºC until used. The SDS-PAGE and the densitometry analysis were done as described for WP. Immunoblotting assays - To determine if the OMP were involved in laminin adhesion, immunoblotting with OMP extracts was done (Doyle 2005). The OMP extracts (3 µL) were dropped onto a nitrocellulose membrane and allowed to dry. The membrane was washed with TBST buffer (10 mM Tris, 150 mM NaCl, 0.1% Tween 20, pH 8.0) and incubated with laminin (20 µg/mL) in blocking buffer (TBST, 5% skim milk, pH 7.4) for 2 h at rt. The membrane was washed (2 x) with TBST buffer for 15 min and incubated with a primary mouse IgG anti-laminin antibody (Santa Cruz; 1:1000) for 1 h at rt. The membrane was washed (2 x) and incubated with a secondary antibody (rabbit anti-mouse conjugated to alkaline phosphatase; Invitrogen; 1:800). Finally, blots were washed (3 x) in TBST and developed. Laminin was used as a positive control. Tests were performed in duplicate. Affinity column - Five hundred microlitres of Affi-Gel® 10 (Bio Rad) was washed 2 x with 0.1 M PBS and mixed immediately with a solution of laminin (1 mg/mL in 0.1 M PBS). The column was blocked with 50 mM ethanolamide pH 7.8 and washed (3 x) with 0.1 M PBS, followed by a wash with 2 M NaCl pH 7.0. OMP extract (1-5 mg/mL) from ATCC strain 43859 was passed several times through the column. After washing the column (3 x) with PBS, a solution of 2 M NaCl pH 7.0 was passed to elute the proteins (Kern & Schotz 1987). After desalting, the samples were subjected to SDS-PAGE as described previously. RESULTS ATCC strain 43859, in both conditions, grew at a comparable rate for 6 h, as measured by optical density. With 8 h and 18 h of incubation, the strain cultured under oxidising condition showed a significant delay in its growth when compared to the strain cultured under reducing condition (p < 0.05). At 24 h, similar behaviour was verified (Fig. 1). It was observed that the strain analysed was capable of adhering to the different ECM components tested (Fig. 2) when the bacterial cells were obtained from cultures kept under either oxidising or reducing conditions after 18 h of growth, with an agglutination titer of at least 10. A distinct capacity to adhere to one of the components tested, laminin, could be detected when the oxidising and reducing conditions were compared. The RR staining demonstrated that in the same population of ATCC strain 43859, grown both under oxidising or reducing conditions, cells were observed with a thick electrondense layer as compared to other cells that were not stained (Fig. 3). The total quantity of polysaccharide, in the sugar assay, expressed under different Eh conditions, confirmed this behaviour. There was not a significant difference (p > 0.05) in the quantity of polysaccharide produced by this strain when both conditions were compared (Fig. 4). A partial hydrolysis of the polysaccharides demonstrated that all antibodies tested were capable of recognising the bacterial extracts, but there was no difference observed between oxidising and reducing conditions. On the other hand, in relation to the crude extract, none of the antibodies except the Bf12 were capable of reacting, and again no difference was observed between the two conditions (Table). The electrophoresis profiles of the WP (Fig. 5A) showed relatively similar patterns when the conditions were compared; however, the differences between the two conditions may be more clearly observed when examining the electrophoresis profiles of the OMPs (Fig. 5B). When comparing the OMP extracts, some variability in the protein expression could reliably be detected. A protein around 49 kDa was observed only under oxidising condition. Five proteins, near 60 kDa, 49 kDa, 39 kDa, 27 kDa and 15 kDa, were expressed more under oxidising than reducing conditions (Fig. 5B). The densitometry of the gel reflected these differences. The immunoblotting analyses confirmed the involvement of the OMP in the adhesion to laminin (Fig. 6). We observed that, when cultured under high (oxidising) Eh, some proteins capable of adhering to laminin were expressed more than when the bacterial cells were cultured under low (reducing) Eh. This result was confirmed when the OMP extracts obtained from cultures kept under oxidising conditions were passed through an affinity column and revealed bands of approximately 60 kDa, 36 kDa, 25 kDa and 15 kDa. For the reducing conditions, only a band of approximately 60 kDa was detected (Fig. 7). DISCUSSION Many events are involved in bacterial infections, and those events can compromise the functionality of a tissue, or result in its destruction. The adherence of microorganisms to the host tissue is the first event and a crucial moment for colonisation. The whole adherence process involves many surface bacterial components, namely, adhesins that can recognise molecules in the host tissue or even components of the ECM, to colonise, invade and spread in the host (Patti et al. 1994). MSCRAMMS have been widely studied and represent an essential factor in the pathogenicity of some microorganisms. In pathogens such as Pseudomonas aeruginosa (Plotkowski et al. 1996), Staphylococcus aureus (Peacock et al. 1999), Yersinia pestis (Lähtennmaki et al. 1998) and Mycobacterium leprae (Marques et al. 2001), MSCRAMMS that bind to laminin have already been identified. It was also reported that B. fragilis may recognise laminin, fibronectin, vitronectin and collagens (Nagy et al. 1994, Eiring et al. 1995). However, no data are available about the role played by such molecular recognition and binding in the infections caused by the species. Recently, Ferreira et al. (2006) have reported the ability of B. fragilis strains to adhere to laminin-1 and concluded that the molecules responsible for the recognition and binding to laminin were in the OMP extracts. In the present study, to evaluate the interference of Eh in B. fragilis adherence to ECM components, we used a simple and rapid method, the latex agglutination assay. The strain used in our study, ATCC 43859, revealed extensive adhesion to laminin when the bacterial cells tested were obtained from cultures kept under oxidising conditions. This strain was also capable of adhering to the other components tested. Some studies have demonstrated that pathogenic bacteria can adhere to molecules such as fibronectin and heparin sulfate to cause systemic infections (Henry-Stanley et al. 2005) and certainly these data related to B. fragilis should be better explored in further investigations. Even though other authors have demonstrated the capacity of B. fragilis to adhere to laminin, this report shows the difference in adherence to this molecule when the strain is grown under oxidising and reducing conditions. The basal membrane, which is rich in laminin, is thin and has an intimate contact with the cytoplasmic membrane of epithelial and endothelial cells (Inou 1989). Although there are at least 11 different isoforms of the laminin molecule, the best studied is the laminin type 1, isolated from EHS, used in our study. The basal membrane represents one of the barriers to B. fragilis establishment and infection, following invasion of the peritoneal cavity and dissemination to other non-intestinal sites. Under normal circumstances, B. fragilis stays in harmony with the host and its growth is controlled. When lesions are formed, its microenvironment can become oxidising and B. fragilis turns into a pathogenic bacterium. Some studies have demonstrated that, when the Eh is altered, it can influence the behaviour of B. fragilis, which becomes more aggressive and invasive in HeLa cells (Goldner et al. 1993). ATCC strain 43859 has a peculiar characteristic; it was isolated from diarrhea and produces the B. fragilis toxin (BFT). This toxin can cause morphological and histological alterations, especially in polarised cells (Sears 2006). Cell polarisation derives from the signals that come from surface, cell-to-cell and cell-ECM activity, forming specialised membrane domains. The adhesion of proteins to the basal membrane and of integrins to the ECM, is essential to cell polarisation (Gumbiner 1996). A subsequent role of BFT and an adhesion to laminin can be speculated and might emerge as an interesting field for further investigations. The interference of Eh in the bacterial growth of ATCC 43859 showed that, under oxidising conditions, there was a significant delay when compared to reducing conditions, but both conditions permitted the bacteria to reach the beginning of the log phase at the same time (after about 6 h). Based on this fact, we decided to compare the behaviour of this strain at a time after 18 h, when the growth curves showed the greatest difference. In fact, the most studied B. fragilis virulence factor is the CPC. This complex has a zwitterion nature (Stingele et al. 2004) and a critical involvement in abscess formation (Coyne et al. 2000). B. fragilis can modulate its surface antigenicity (phase dependent) and it has at least eight distinct polysaccharides (PSA to PSH) (Krinos et al. 2001). It is well documented that the CPC is involved in abscess formation (Stingele et al. 2004). This capsule modulation (Krinus et al. 2001) may be used as a strategy to help the bacteria evade the host immune system and at the same time to colonise the host. The TEM of ATCC strain 43859, using ruthenium red, revealed a very diverse population of cells, some with a thicker polysaccharide layer and others with none. This heterogeneity was verified in cultures kept in oxidising and reducing conditions. Patrick et al. (1999) have described that, in the same culture of B. fragilis, it is possible to find cells expressing at least three different kinds of capsule that could be visualised under TEM: thick, thin and an electron dense layer (Patrick 1993). The qualitative analysis of the polysaccharides confirmed this result. The two strains tested, ATCC 43859 and ATCC 25285, were recognised by antibodies after partial hydrolysis, and there was no difference between one condition and another. When the crude extract of ATCC strain 25285 was used, the Bf12 antibody recognised it, but still with no difference between the two Eh conditions. Our results suggest that polysaccharide expression is neither influenced by the Eh nor involved in the recognition of the laminin. SDS-PAGE of the WP of the ATCC strain 43859 did not show obvious differences between oxidising and reducing conditions. In contrast, electrophoretic profiles of the OMP showed differences between the extracts obtained from cells cultured in the two Eh conditions. Immunoblotting with OMP extracts revealed a difference in the recognition of laminin when the proteins were extracted from different culture conditions, oxidising and reducing. The laminin affinity column detected proteins of approximately 60 kDa, 36 kDa, 25 kDa and 15 kDa extracted from oxidised conditions. For the reduced conditions, only one band, at 60 kDa, was observed. Ferreira et al. (2006) have already demonstrated the involvement of the OMPs of B. fragilis in the adherence of laminin, and resident amino acid sequences inhibited this recognition. Our results suggest that structures present on the B. fragilis surface can be influenced by Eh; moreover, the adhesins responsible for laminin recognition are manifested in the OMP extracts, and further investigations are being done to confirm this. ACKNOWLEDGEMENTS To Prof. J. Smalley and Mr. A. Birss (University of Liverpool), for the use of an anaerobic chamber, to Dr. Sheila Patrick and L. McCoy (Queen's University), for monoclonal antibodies, and Joaquim dos Santos, for technical support. REFERENCES
Copyright 2008 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc08124f6.jpg] [oc08124f5.jpg] [oc08124f2.jpg] [oc08124t1.jpg] [oc08124f7.jpg] [oc08124f1.jpg] [oc08124f3.jpg] [oc08124f4.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}