 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 103, No. 7, November, 2008, pp. 736-740 SHORT COMMUNICATIONS Genetic divergence between two sympatric species of the Lutzomyia longipalpis complex in the paralytic gene, a locus associated with insecticide resistance and lovesong production RMMA LinsI; NA SouzaII; AA PeixotoI, + ILaboratório
de Biologia Molecular de Insetos Financial support: Howard Hughes Medical Institute, CNPq, Fiocruz (PAPES IV) Received 27 May
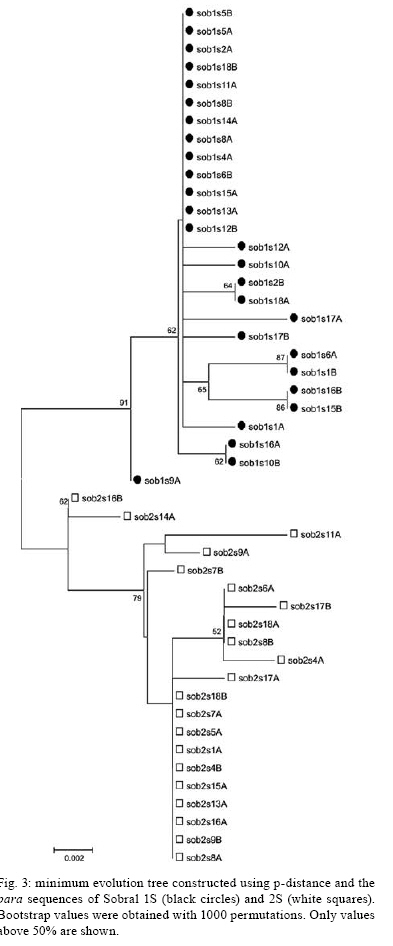
2008 Code Number: oc08133 ABSTRACT The sandfly Lutzomyia longipalpis s.l. is the main vector of American Visceral Leishmaniasis. L. longipalpis s.l. is a species complex but until recently the existence of cryptic sibling species among Brazilian populations was a controversial issue. A fragment of paralytic (para), a voltage dependent sodium channel gene associated with insecticide resistance and courtship song production in Drosophila, was isolated and used as a molecular marker to study the divergence between two sympatric siblings of the L. longipalpis complex from Sobral, Brazil. The results revealed para as the first single locus DNA marker presenting fixed differences between the two species in this locality. In addition, two low frequency amino-acid changes in an otherwise very conserved region of the channel were observed, raising the possibility that it might be associated with incipient resistance in this vector. To the best of our knowledge, the present study represents the first population genetics analysis of insecticide resistance genes in this important leishmaniasis vector. Key words: Lutzomyia longipalpis - Drosophila - insecticide resistance - para - Leishmaniasis - sandflies - sympatric species The sandfly Lutzomyia longipalpis s.l. (Lutz & Neiva 1912) (Diptera: Psychodidae: Phlebotominae) is the main vector of Leishmania chagasi (= Leishmania infantum), the causative agent of American Visceral Leishmaniasis. Because of its importance as a vector, several studies have been carried out to verify the taxonomic status of L. longipalpis and there is now a large amount of data indicating that this sandfly is a species complex (reviewed in Bauzer et al. 2007, Maingon et al. 2008). Although the current evidence clearly indicates the existence of sibling species among Brazilian populations, the issue was a controversial one until recently (Bauzer et al. 2007, Maingon et al. 2008). The best evidence for the existence of sibling species in Brazil can be found in Sobral (state of Ceará). In this locality males of two sympatric species can be differentiated by the presence of one (1S) or two (2S) abdominal spots (Ward et al. 1988, Souza et al. 2008), produce different types of pheromones and copulatory courtship songs (Souza et al. 2004, Hamilton et al. 2005) and show a significant genetic divergence with different molecular markers (Bauzer et al. 2002a, Maingon et al. 2003, Bottecchia et al. 2004). Genes controlling the production of acoustic signals during courtship are interesting molecular markers to study the speciation process since these "lovesongs" are an important aspect of sexual behaviour and reproductive isolation in many insect species. A number of genes controlling the courtship song in Drosophila melanogaster have been identified (Gleason 2005) and two of these, period and cacophony, have already been used in studies of the L. longipalpis complex in Brazil (Bauzer et al. 2002a,b, Bottecchia et al. 2004). One of the genes controlling the lovesong in D. melanogaster is paralytic (para) (Peixoto & Hall 1998). Interestingly, mutations in para orthologues are also associated with insecticide resistance in a number of organisms (Hemingway & Ranson 2000, Hemingway et al. 2004). This gene codes for the subunit of a voltage-dependent sodium channel (Loughney et al. 1989, Littleton & Ganetzky 2000) and its homologues in other insects are the site of a number of mutations associated with resistance to DDT and pyrethroids, including the well known kdr mutation (Milani 1956, Loughney et al. 1989, Pittendrigh et al. 1997). Insecticide resistance in vectors of tropical diseases is a major public health problem. Nevertheless, in many vector species such as Neotropical sandflies the occurrence of insecticide resistance is still poorly studied. The use of resistance genes as molecular markers offers an attractive approach to investigate the rise and spread of insecticide resistance mutations in populations of vector species. Interestingly some authors speculate whether insecticide resistance is a consequence of insecticide introduction or if these mutations arose before it (Hartley et al. 2006, ffrench-Constant 2007). It has been shown that mutations conferring resistance to Malathion in blowflies of the genus Lucilia were already present before the introduction of this insecticide (Hartley et al. 2006). This implies that species carrying such alleles are pre-adapted to insecticide exposition. The occurrence of insecticide resistance in L. longipalpis has not yet been reported. There are only a handful of studies on this subject in sandflies (e.g. Rahman et al. 1982, Mukhopadhyay et al. 1987, 1996, Tetreault et al. 2001, Surendran et al. 2005, Coutinho-Abreu et al. 2007) and further research is needed, particularly in Neotropical species. Therefore, the isolation of L. longipalpis molecular markers homologous to genes controlling insecticide resistance in other insects might help to elucidate this question. We have used a degenerate primer PCR approach to clone a L. longipalpis fragment homologous to the para gene of Drosophila. The primers were designed to amplify the region encoding the IIS6 transmembrane segment of the channel known to contain insecticide resistant mutations in other insects. A population genetic analysis was also carried out using the two sympatric species of the locality of Sobral. A sample of L. longipalpis from a laboratory colony originated from sandflies collected at Lapinha cave (state of Minas Gerais, Brazil) was used to isolate the initial fragment. The samples from Sobral were the same analysed by Bauzer et al. (2002a) and Bottecchia et al. (2004). For the initial fragment of the para gene, mRNA was isolated with QuickPrepTM Micro mRNA purification kit (Amersham Biosciences) and cDNA was synthesized using the First-Strand cDNA synthesis kit (Amersham Biosciences). Initially, touchdown PCR was performed in 40 µL using Promega reagents with various cycling conditions and the degenerate primers: 5paraIIdegC (5´ - TGGAAYTTYACNGAYTT - 3´) and 3paraIIdegB (5´ - TTRTTNGTRTCRTTRTC - 3´). Degenerate primers were designed based on conserved regions identified by alignment of the protein sequences encoded by para orthologues in different insect species. Based on the initial fragment generated, specific primers were designed. For the population genetics analyses of the two Sobral siblings, genomic DNA of individual flies were isolated according to Jowett (1998) and the PCR Master Mix (Promega) was used to perform reactions according to the following cycling conditions: 95ºC for 3 min; 35 cycles at 95ºC for 30 s, 60ºC for 30 s and 72ºC for 30 s. Specific primers used were: 5llpara2 (5´ - ACGGACTTCATGCATTCATTC - 3´) and 3llpara1 (5´ - TGGTGCTGATAAACTTGACG - 3´). PCR products were purified using the Wizard SV Gel and PCR Clean-up System (Promega) and cloned using the pMOSBlue Blunt Ended Cloning Kit (Amersham Biosciences). Plasmidial DNA was isolated using Flexiprep Kit (Amersham Biosciences) or using 96 wells microplates and the alcaline lysis method followed by filtration in Millipore Multiscreen filter plates. DNA sequencing was carried out in ABI377 and ABI 3730 Sequencers using the Big Dye 3.1 Kit (Applied Biosystems). Homology to the Drosophila para gene was confirmed by comparison with databanks using BlastX (http://www.ncbi.nlm.nih.gov). Fragments of the para gene of L. longipalpis from Sobral 1S and 2S were initially processed using GCG program (Wisconsin Package Version 9.1, Genetic Computer Group, Madison, Wisconsin, USA) before the population genetics analyses. A minimum of three and maximum of eight sequences per individual were aligned to obtain one or, when possible, two consensus sequences corresponding to the two alleles, named as A and B. Polymorphism and population structure analyses were carried out using both DnaSP 4.10.4 (Rozas et al. 2003) and Proseq 2.9 (Filatov & Charlesworth 1999) programs. Phylogenetic analysis was performed using Mega 3.1 (Kumar et al. 2004). Sequences were submitted to the GenBank (accession numbers DQ914434; EU746318-EU746365). Fig. 1 shows the amino acid sequence encoded by the initial fragment obtained from L. longipalpis aligned with the respective region of the para protein of D. melanogaster. This region is highly conserved with only one amino acid substitution (Y→L) in L. longipalpis. The intron position is also conserved. We have generated 27 consensus sequences of Sobral 1S and 21 sequences of Sobral 2S, where 376 sites were analyzed and 23 of them (6.1%) were variable. Fig. 2 shows the alignment of polymorphic sites observed in the different haplotypes generated. Most of the variation found is within the intron which also shows a few insertion deletion events. We also have found one synonymous (position 19) and two non-synonymous substitutions (position 114, N→K, Sob2S17B and position 369, N→K, Sob2S14A). The positions of these two non-synonymous substitutions are underlined in the alignment shown in Fig. 1. Compared to the other two lovesong genes analysed in Sobral (Bauzer et al. 2002a, Bottecchia et al. 2004), the level of polymorphism in para is low ( π= 0.0037 andθ= 0.0091 for Sobral 1S and π= 0.0045 and θ= 0.0075 for Sobral 2S). FST analysis showed a very high and significant differentiation between Sobral 1S and Sobral 2S (FST = 0.7852; Nm = 0.07, p < 0.0001). In addition, three fixed differences were observed between the two siblings. Fig. 3 shows a minimum evolution tree of the sequences of the two sympatric siblings. This tree shows a clear separation between the sequences of the two species with a high bootstrap value of 91%. The tree obtained shows a similar topology to that using the period gene (Bauzer et al. 2002a). Although insecticide resistance has not yet been demonstrated in L. longipalpis there are some indications of its occurrence in this species (Falcão et al. 1988, Mazzarri et al. 1997). The fragment we isolated may provide a useful molecular marker for the screening of putative insecticide resistance mutations in populations of this vector. In fact, although the non-synonymous substitutions observed in Sobral 2S were not associated with insecticide resistance in other species, these amino-acid changes occur in an otherwise very conserved region of the channel, raising the possibility that it might be associated with incipient resistance in this vector. Therefore, it might be interesting to screen other populations, especially those under current treatment with pyrethroids, for the presence of these mutations. As mentioned above, L. longipalpis is a species complex (Bauzer et al. 2007, Maingon et al. 2008) and males of different sibling species produced very distinct copulatory courtship songs (Souza et al. 2004). In addition to its role in insecticide resistance, para is also involved in the control of the courtship song in Drosophila (Peixoto & Hall 1998). Therefore para is a very good candidate as an additional molecular marker for further studies in this species complex. Although fixed differences in isoenzyme loci have been reported (e.g., Lanzaro et al. 1993, Lampo et al. 1999) and microsatellites have been used successfully in the genetic identification of L. longipalpis siblings (Maingon et al. 2003), para is the first single locus DNA marker presenting fixed differences between the two sympatric species from Sobral. This might allow the development of a field friendly agarose assay to differentiate the siblings. Therefore this locus might be especially useful as a tool to species identification of females in field studies aimed at comparing the infection rates and vectorial capacity between these two sympatric sibling species of the L. longipalpis complex. REFERENCES
Copyright 2008 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc08133f3.jpg] [oc08133f2.jpg] [oc08133f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}