 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 104, No. 1, February, 2009, pp. 48-55 Biological activity of neosergeolide and isobrucein B (and two semi-synthetic derivatives) isolated from the Amazonian medicinal plant Picrolemma sprucei (Simaroubaceae) Ellen CC SilvaI, II; Bruno C CavalcantiIV; Rodrigo CN AmorimI, II; Jorcilene F LucenaI, II; Dulcimar S QuadrosI, V; Wanderli P TadeiIII; Raquel C MontenegroIV; Letícia V Costa-LotufoIV; Cláudia PessoaIV; Manoel O MoraesIV; Rita CS NunomuraI; Sergio M NunomuraI; Marcia RS MeloI; Valter F de Andrade-NetoVII; Luiz Francisco R SilvaII, VI; Pedro Paulo R VieiraVI; Adrian M PohlitI, + ILaboratório
de Princípios Ativos da Amazônia, Coordenação de
Pesquisas em Produtos Naturais Financial support: CNPq/PPG-7 (557106/2005-2, 563892/2005-6), CNPq/PNOPG (520354/1999-0, 550260/2001-3), Bioamazonia-Basa-Fepad Contract Received 9 June
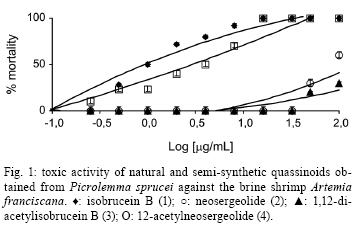
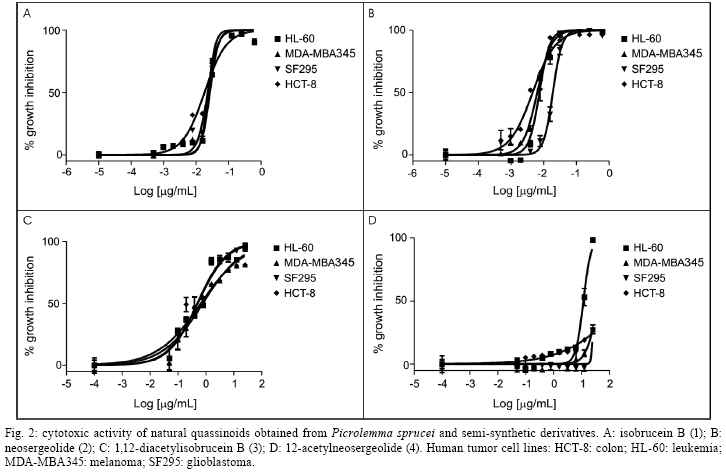
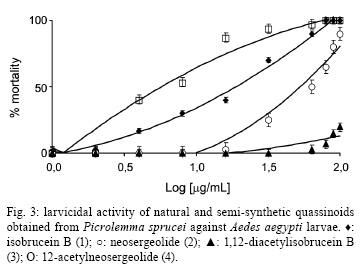
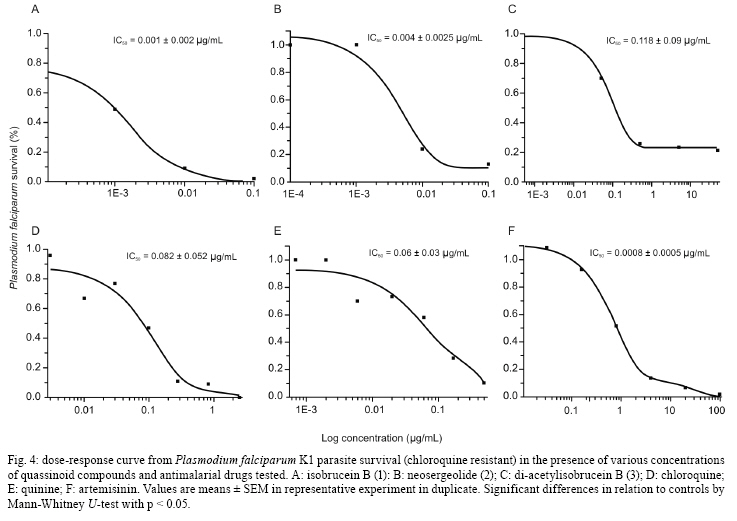
2008 Code Number: oc09008 ABSTRACT In the present study, in vitro techniques were used to investigate a range of biological activities of known natural quassinoids isobrucein B (1) and neosergeolide (2), known semi-synthetic derivative 1,12-diacetylisobrucein B (3), and a new semi-synthetic derivative, 12-acetylneosergeolide (4). These compounds were evaluated for general toxicity toward the brine shrimp species Artemia franciscana, cytotoxicity toward human tumour cells, larvicidal activity toward the dengue fever mosquito vector Aedes aegypti, haemolytic activity in mouse erythrocytes and antimalarial activity against the human malaria parasite Plasmodium falciparum. Compounds 1 and 2 exhibited the greatest cytotoxicity against all the tumor cells tested (IC50 = 5-27 µg/L) and against multidrug-resistant P. falciparum K1 strain (IC50 = 1.0-4.0 g/L) and 3 was only cytotoxic toward the leukaemia HL-60 strain (IC50 = 11.8 µg/L). Quassinoids 1 and 2 (LC50 = 3.2-4.4 mg/L) displayed greater lethality than derivative 4 (LC50 = 75.0 mg/L) toward A. aegypti larvae, while derivative 3 was inactive. These results suggest a novel application for these natural quassinoids as larvicides. The toxicity toward A. franciscana could be correlated with the activity in several biological models, a finding that is in agreement with the literature. Importantly, none of the studied compounds exhibited in vitro haemolytic activity, suggesting specificity of the observed cytotoxic effects. This study reveals the biological potential of quassinoids 1 and 2 and to a lesser extent their semi-synthetic derivatives for their in vitro antimalarial and cytotoxic activities. Key words: neosergeolide - isobrucein B - 12-acetylneosergeolide - 1,12-diacetylisobrucein B - cytotoxicity - antimalarial - larvicide The global malaria situation has worsened in the last two decades with an estimated 350-500 million new cases and more than one million annual deaths attributed to this disease, 90% of which have been recorded in Africa (WHO 2005, CDC 2008). Global efforts toward controlling malaria are greatly challenged by the increasing spread of antimalarial drug resistance and the use of ineffective antimalarials is considered partially responsible for the difficulties in reducing malaria morbidity and mortality (Vastergaard & Ringwald 2007). New drugs are therefore needed to treat malaria. Interest in plants as sources of new antimalarials has been stimulated by the isolation of artemisinin from Artemisia annua. This compound is active against drug-resistant Plasmodium falciparum, the most lethal human malaria parasite species (van Agtmael et al. 1999). The Brazilian Amazon, where human malaria is endemic, is one of the richest sources of plants with potential pharmacological activity. Plants are frequently used to treat the disease as infusions or as additives in alcoholic beverages (Milliken 1997). Picrolemma pseudocoffea Ducke (pseudonym for Picrolemma sprucei Hook. f.) is one of a number of plant species known in the Brazilian Amazon as caferana and is commonly used for the treatment of malaria throughout the Amazon region (Grenand et al. 1987, Duke & Vasquez 1994, Milliken 1997). In the investigation of the biological activity of plant extracts and natural products isolated from plants, the brine shrimp assay is a valuable tool for establishing general toxicity and cytotoxicity parameters. This assay consists of exposing brine shrimp larvae to plant extract in saline solution and larval mortality is evaluated after one day. A very positive correlation between the lethality to brine shrimp and cytotoxicity has been established by researchers working on the development of new anti-cancer drugs from plants at the National Cancer Institute in the United States (Anderson et al. 1991). This correlation is considered so good that lethality toward brine shrimp is recommended by these authors as an effective pre-screen for existing in vitro cytotoxicity and antitumor assays (Anderson et al. 1991, Quignard et al. 2003). Invasive mosquitoes are economic and sanitary concerns, especially in Europe and America (Boyer et al. 2006). Among the mosquito vectors of most concern is Aedes aegypti (Diptera: Culicidae) which originated in Africa, has spread around the world and is a vector of yellow and hemorrhagic dengue fever. The present resurgence of these diseases is due to the higher number of breeding places for these mosquitoes in today's throwaway society (Kiran et al. 2006). Additionally, temephos, a synthetic organophosphate compound widely used as insecticide and larvicide in Brazil presents related resistance cases and therefore a loss of overall efficacy (Lima et al. 2006) which is compounded in tropical countries having high annual rainfalls by the need for as little as 1-2 mL of clean water for this species to carry out the aquatic portion of its life cycle, from egg through larval phase to emergent imago (adult). For these reasons and despite significant advances in techniques used for its control during recent decades, A. aegypti and other mosquitoes continue to pose serious public health problems (Gusmão et al. 2002). Importantly, co-evolution has equipped plants with a plethora of chemical defences against insect predators. As such, mankind has used plants or plant extracts to control insects since ancient times (Balandrin 1985, Sukamar et al. 1991). In previous work, we have shown the larvicidal activity against A. aegypti of P. sprucei stem and root methanol extracts (Pohlit et al. 2004). Quassinoid is the name given to any of a number of bitter substances found exclusively in the Simaroubaceae family (Polonsky 1973). Chemically, quassinoids are degraded triterpene compounds which are frequently highly oxygenated. Many quassinoids exhibit a wide range of biological activities in vitro and/or in vivo, including antitumor, antimalarial, antiviral, anti-inflammatory, antifeedant, insecticidal, amoebicidal, antiulcer and herbicidal activities. Bruceantin, simalikalactone D, quassin, brusatol and glaucarubinone are some of the most well-studied quassinoids and exhibit a wide range of biological activities (Guo et al. 2005). Quassinoid compounds isobrucein B (1) and neosergeolide (2) have been isolated previously from the stems and roots of P. sprucei (Moretti et al. 1982, Schpector et al. 1994) and isobrucein B was also obtained from the leaf extract of this same plant species (supplementary data) (Polonsky et al. 1984). Our group has recently demonstrated the important antimalarial activity of 2 (Andrade-Neto et al. 2007), the antihelminthic activity of 1 and 2 (Nunomura et al. 2006) and has written a review in which the noteworthy antileukemic, antifeedant and leishmanicidal activities of 1 were discussed (Amorim & Pohlit 2006). Kupchan et al. (1975) prepared the semi-synthetic quassinoid 1,12-diacetylisobrucein B (3) by acetylation of 1, but no evaluation of its biological activity has been published to date. In general, semi-synthetic modification of natural quassinoids is thought to be a useful tool for the discovery of new therapeutic drugs and quassinoid derivatives with other potential uses and could lead to derivatives with increased therapeutic indices, including significantly increased pharmacological activity and/or decreased toxicity. Herein, in vitro methods are used to investigate the general toxicity in the brine shrimp assay towards Artemia franciscana, the cytotoxicity toward human tumor cells, the larvicidal activity in A. aegypti, and the haemolytic and in vitro antimalarial activity against P. falciparum of known isolated quassinoids (1 and 2) and semi-synthetic quassinoid derivative 3, as well as a new semi-synthetic derivative, 12-acetylneosergeolide (4). MATERIALS AND METHODS Plant materials - Collection was performed in Silves, state of Amazonas (AM), Brazil, in April 2003. Voucher specimens have been deposited at the Universidade Federal do Amazonas Herbarium (Silva 5729 & 5730). Identification was performed by Dr. Wayt Thomas as P. sprucei (Wayt Thomas, personal communication). Isolation and identification of quassinoids - The procedure used for the isolation of quassinoids isobrucein B (1) and neosergeolide (2) from the roots and stems of P. sprucei has been described in a previous publication (Andrade Neto et al. 2007). Spectrometric measurements - Nuclear magnetic resonance (NMR) spectra were obtained on a Varian Unity Inova model, using VnmrJ software. Positive ion electrospray ionization accurate mass spectra [(+)-ESI-MS] were obtained on a Bruker-Daltronics UltrOTof apparatus using MeOH/H2O/HCO2H as an infusion solvent system. Melting points were determined on a Marconi MA-381 apparatus and are uncorrected. Fourier-transform infrared spectra were obtained on a BOMEM FTLA-2000-104 and UV spectra were obtained on a Femto 800 XI apparatus. 1,12-diacetylisobrucein B - Treatment of compound 1 (50.6 mg, 105.4 µmol) with pyridine (3.3 mL) and acetic anhydride (3.3 mL) in accordance with Kupchan et al. (1975) yielded 1,12-diacetyl derivative 3 (50.7 mg, 85.2%); mp 240-245°C [264-267°C (Kupchan et al. 1975)]; IR (KBr) 3447 (O-H), 2919-2950 (C-H), 1734-1750 (C = O), 1647 (C-2, C = O) cm-1 [(KBr) 3534, 1745, 1681, 1370, 1230, 1036 cm-1 (Kupchan et al. 1975)]; UVmax (MeOH) 241.0 nm. [UVmax (EtOH) 238 nm (Kupchan et al. 1975)]; 1H NMR (500 MHz, CDCl3) δ 6.06 (s, H-3), 5.38 (s, H-1), 5.26 (s, H-12), 4.79 (d, H-30s, J = 8.3), 4.74 (s, H-7), 4.10 (d, H-11, J = 4.1), 3.79 (s, H-5'), 3.77 (d, H-30a, J = 8.3), 3.25 (d, H-14, J = 11.6), 3.03 (d, H-5, J = 13.0), 2.42 (ddd, H-6e, J = 13, 2.0, 2.0), 2.23 (s, H-9') 2.18-2.25 (m, H-9), 2.10 (s, H-2'), 2.01 (s, H-7'), 1.94 (s, H-29), 1.83 (ddd, H-6a, J = 13.0, 13.0, 2.0), 1.39 (s, H-19) [(100 MHz, CDCl3) δ 5.95 (br s, H-3), 5.15 (d, H-15, J = 15), 3.69 (s, H-5'), 2.16 (s, H-9'), 2.03 (s, H-2'), 1.95 (s, H-7'), 1.88 (s, H-29), 1.14 (s, H-19) Kupchan et al. 1975)]; 13C NMR (125 MHz, CDCl3) δ 190.57 (C-2), 170.34 (C-8'), 169.41 (C-18), 168.85 (C-1'), 166.64 (C-16), 159.96 (C-4), 126.44 (C-3), 83.84 (C-1), 82.70 (C-7), 80.36 (C-13), 74.68 (C-12), 73.92 (C-30), 71.23 (C-11), 53.10 (C-5'), 46.13 (C-10), 45.51 (C-8), 43.5 (C-5), 42.52 (C-9), 28.04 (C-6), 22.57 (C-29), 21.04 (C-9'), 20.93 (C-7'), 20.50 (C-2'), 12.24 (C-19); ESI-tof-MS m/z 587.1775 [M+Na]+ (exact mass for C27H32O13Na+ 587.1735) [EI-MS m/z 564 (M+), 522, 504, 489, 135, 95, 91, 60, 43 (Kupchan et al. 1975)]. 12-acetylneosergeolide - Neosergeolide (2, 16.4 mg, 32.5 mol), pyridine (1 mL), and acetic anhydride (1 mL) were magnetically stirred for four days at rt. After complete evaporation, the residue was dissolved in methanol (1 mL) resulting in the formation of a precipitate. The precipitate was dissolved in methylene chloride-methanol (1:1) and then submitted to preparative thin-layer chromatography (2% isopropyl alcohol in methylene chloride) which resulted in the isolation of 12-acetylneosergeolide as needles (4, 15.1 mg, 85.1%); (supplementary data) mp 214.6°C (decomp.); IR (KBr) 3535 (O-H), 2983-2971 (C-H), 1753 (C-18, C = O), 1679 (C-16, C = O), 1586 (C1'-4, C = O), 1209-1238 (C-O) cm-1; UVmax (MeOH) 279.0 nm; 1H NMR (CDCl3 + CD3OD) δ 5.96 (s, H-4'), 5.55 (t, H-3, J = 2.8), 5.11 (s, H-12), 4.65 (d, H-30s, J = 8.3), 4.57 (s, H-7), 4.19 (d, H-11, J = 3.0), 3.63 (d, H-30a, J = 8.3), 3.60 (s, H-5'), 3.09 (dl, H-14, J = 12.5), 2.20-2.24 (m, H-4), 2.05-2.10 (m, H-9 + H-6e), 1.90 (s, H-2'), 1.82 (s, H-7'), 1.50-1.65 (m, H-5 + H-6a) 1.39 (s, H-19), 1.00 (s, H-29); 13C NMR (CDCl3) δ 172.3 (C-6'), 170.9 (C-3'), 169.8 (C-18), 169.2 (C-1'), 167.2 (C-16), 162.5 (C-1), 148.5 (C-2), 116.3 (C-3), 112.9 (C-4'), 83.4 (C-7), 79.9 (C-13), 75.3 (C-12), 73.5 (C-30), 70.7 (C-11), 52.7 (C-5'), 49.1 (C-14), 46.0 (C-8), 44.9 (C-5), 41.8 (C-10), 39.3 (C-9), 30.7 (C-4), 28.4 (C-6), 20.5 (C-7'), 20.0 (C-2'), 19.2 (C-29), 17.8 (C-19); ESI-tof-MS m/z 547.1810 [M+H]+ (calculated exact mass for C27H31O12+ 547.1810). Brine shrimp assay - The lethality of quassinoids toward nauplii (larvae) of the brine shrimp species A. franciscana Kellogg was evaluated according to established procedures (Meyer et al. 1982, Anderson et al. 1991, Mongelli et al. 1996, Parra et al. 2001, Quignard et al. 2003). Quassinoids and derivatives were separately dissolved and diluted in DMSO to provided sample solutions having 10 concentrations ranging from 1000-0.25 µg/mL. The co-solvent DMSO had final well concentrations of < 1% in all experiments and controls. For the brine shrimp assay, wells of 24-well microtitre plates were pre-filled with 0.8 mL saline solution. Next, 10 two-day old nauplii (2nd instar larvae) in a minimum volume of saline solution (ca. 0.2 mL), 10 µL of each quassinoid dissolved in DMSO and a stock saline solution were added, which provided a final volume of 1.0 mL in each well (Mosmann 1983). Each compound was tested in three independent assays, each in triplicate. The plates were then allowed to stand in the absence of direct light (to avoid possible false positive results due to the generation of phototoxic compounds) at rt and mortality was evaluated after 24 h. The half-maximal inhibitory (IC50) responses, as compared to the drug-free controls, were estimated by Probit analysis using Microsoft Excel® software. MTT assay - Quassinoids and derivatives were tested for cytotoxicity in four human tumour cell lines: SF295 (glioblastoma), MDA-MBA345 (melanoma), HCT-8 (colon) and HL-60 (leukemia) (Children's Mercy Hospital, Kansas City, MO, USA). Cells were cultured in RPMI-1640 medium supplemented with 10% foetal bovine serum, 2 mM glutamine, 100 µg/mL streptomycin and 100 U/mL penicillin and incubated at 37°C under a 5% CO2 atmosphere. Cells were then plated in 96-well plates (105 cells/well for adherent cells or 0.5 x 105 cells/well for suspended cells in 100 µL of medium). After 24 h, compounds (25.0 to 0.001 µg/mL in 17 dilutions) dissolved in 1% DMSO were added to each well and incubated for 72 h. Doxorubicin (0.01-0.58 µg/mL) was used as positive control. Tumor cell growth was quantified by the ability of living cells to reduce the yellow dye 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2H -tetrazolium bromide to a purple formazan product (Mosmann 1983). The drug effect was quantified as the percentage of the absorbance of reduced dye at 595 nm in relation to control wells. The IC50 responses as compared to the drug-free controls were estimated by interpolation using Prism4® software. Larvicidal activity in A. aegypti - Larvicidal activity of quassinoids toward A. aegypti was evaluated using the method described in Pohlit et al. (2004). A. aegypti larvae were obtained by incubation of eggs from a laboratory colony (originated from an AM field strain) at Coordenação de Pesquisas em Ciências da Saúde, Instituto Nacional de Pesquisas da Amazônia, in tap water. Quassinoids and derivatives (0.1 to 100 µg/mL in 11 dilutions) were dissolved in DMSO, which was used as the co-solvent at final well concentrations of < 1% in all experiments and controls. Temephos (0.01-0.58 µg/mL) was used as positive control. Negative controls contained 1% DMSO instead of sample solution. Each experiment and controls were performed in three independent assays, each in triplicate, and compared with controls after 24 h and 48 h at 26-27°C. The IC50 responses as compared to the drug-free controls were estimated by Probit analysis using Microsoft Excel® software. Parasite culture and in vitro antimalarial tests - Chloroquine, pyrimethamine and cycloguanil resistant P. falciparum strain K1 was acquired from MR4 (Malaria Research and Reference Reagent Resource Centre, Manassas, Virginia, USA) and was used in all in vitro tests. Parasites were maintained in continuous culture in A+ human erythrocytes using RPMI medium supplemented with 10% human serum, as described by Trager and Jensen (1976). The antiparasitic effect of the compounds was measured by growth inhibition percentages as described by Carvalho and Krettli (1991). Trophozoite-stages in sorbitol-synchronized blood (Lambros & Vanderberg 1979) were cultured at 1-2% parasitaemia and 2.5% haematocrit and then incubated with compounds (maximum 1 mg/mL in serial dilutions) diluted with a 0.02% final concentration of DMSO in the culture medium (RPMI 1640) for a total of 48 h at 37ºC. Positive controls contained reference antimalarial drugs (chloroquine, quinine and artemisinin) in standard concentrations (Rieckmann et al. 1978, WHO 2001) and were used for each experiment. The stock solutions were further diluted with complete medium (RPMI 1640 plus 10% human serum) to each of the concentrations used (100-0.0001 µg/mL in 7 dilutions). The IC50 responses as compared to the drug-free controls were estimated by interpolation using Microcal Origin® software. Each duplicate experiment was repeated three times and blood smears were read blind (Andrade-Neto et al. 2007). Haemolytic assay - This test was performed in 96-well plates using a 2% mouse erythrocyte suspension in 0.85% NaCl containing 10 mM CaCl2, following the method described by Costa-Lotufo et al. (2002). Compounds were tested at concentrations ranging from 1.5-200 µg/mL. 0.1% Triton X-100 (in 0.85% saline) was used as a positive control. After incubation at rt for 1 h and centrifugation, the supernatant was removed and the liberated haemoglobin was measured spectrophotometrically as the absorbance at 540 nm. Statistical analysis - Data are presented as mean value ± SEM. The IC50 or EC50 values and their 95% confidence intervals (CI 95%) were obtained by nonlinear regression using the GRAPHPAD program (Intuitive Software for Science, San Diego, CA). LC50 values for the brine shrimp assay were obtained from 24 h larvae counts using the probit analysis method described by Finney (1971). RESULTS AND DISCUSSION Known and new quassinoid derivatives 3 and 4, respectively, were prepared using simple acetylation-purification procedures and the ESI-MS, IR and NMR spectra were consistent with the proposed structures. Importantly, spectroscopic evidence supports the regiochemical outcome of the acetylation, which could, a priori, have occurred at either or both the C-11 or C-12 hydroxyl group. NMR-based evidence suggests that the C-12 hydroxyl group undergoes acetylation (together with the C-1 hydroxyl in the case of isobrucein B acetylation) preferentially over the C-11 hydroxyl group. Namely, the signal at δ 5.11 in the 1H-NMR spectrum of 4 was assigned to H-12. In the Heteronuclear Single Quantum Coherence spectrum, this signal had a 13C-1H correlation with δ 75.31 (C-12). The H-12 signal of 4 was shifted approximately 1 ppm to lower field relative to H-12 (δ 4.23) of the precursor 2. This downfield shift is evidence for the deshielding caused by the electronegative acetate group on C-12 in derivative 4. Furthermore, the assignment of H-12 (δ 5.11) is supported by long-range 13C-1H correlations with C-13 (δ 79.9) and C-18 (δ 169.8) in the Heteronuclear Multiple Bond Correlation spectrum as well as correlation with neighbouring H-11 (δ 4.19) in the 1H-1H Correlation Spectroscopy spectrum of 4. The brine shrimp assay is a simple, inexpensive method which allows for the evaluation of general toxicity and is considered a preliminary assay in the study of extracts and secondary metabolites for cytotoxic and antitumor activity. LC50 values in the brine shrimp assay correlate well with other in vitro tests such as those for antitumor activity, cytotoxicity (Meyer et al. 1982, Munro et al. 1987, Siqueira et al. 2001) and antineoplastic activity (Badaway & Kappe 1997) and also for in vivo toxicity (LD50) of extracts administered by gavage in Swiss albino mice (Parra et al. 2001) and pesticidal (Fatope et al. 1993), larvicidal, fungicidal, molluscicidal (Cepleanu et al. 1994), antimalarial (Pérez et al. 1997), insecticidal (Oberlies et al. 1998) and antifeedant activities (Labbe et al. 1993). Quassinoids 1 and 2 were considered very toxic toward A. franciscana, while 3 showed low toxicity (Fig. 1, Table). The same was observed for the MTT cytotoxicity tests, where 1 and 2 exhibited IC50 values ranging from 0.005-0.027 g/mL for all tumor cell lines, while 3 and 4 exhibited IC50 varying from 0.51 to > 25 µg/mL (Fig. 2, Table). In preliminary work, it was shown that at a concentration of 500 µg/mL, P. sprucei stem and root methanol extracts caused 57% and 74% mortality in A. aegypti larvae, respectively (Pohlit et al. 2004). In the present study, since quassinoids 1 and 2 are present in methanol and other polar extracts, the potential larvicidal activity of 1 and 2, as well as their acetylated derivatives 3 and 4, respectively, were investigated. The results showed that 1, 2 and 4 were active against A. aegypti third instar larvae (Fig. 3); compound 3 showed no activity at a dose of 100 µg/mL. As such, quassinoids 1 and 2 present in P. sprucei could be partially responsible for the larvicidal activity exhibited by extracts of this plant. The synthetic compound temephos was very lethal toward A. aegypti larvae (IC50 = 0.025 µg/mL). Quassinoids and their derivatives were less active than temephos, but could be important models for the further development of new larvicides. There is no data on the effectiveness of these compounds in a field setting, nor is it known whether toxicity toward other organisms could be a problem in a field application. Notably, due to reported resistance cases and concerns associated with permanence of organophosphates in the environment over extended periods of time, there is a need to find substitutes for or alternatives to organophosphate insecticides (Lima et al. 2006). Significant inhibition of the human malaria parasite P. falciparum in vitro was exhibited by quassinoids 1, 2 and derivative 4. Ranked on a molar scale, the order of increasing inhibition of parasites was 3 < 4 < 2 < 1 (IC50 = 34.91, 0.216, 0.008 and 0.002 µM, respectively) (Fig. 4). Chloroquine, quinine and artemisinin displayed IC50 values of 0.89, 0.012 and 0.002 µM, respectively. As such, neosergeolide (2) and isobruceín B (1) presented the highest activity, in the same range of activity as the clinically important drug standards that were used as controls. Despite moderate parasite inhibition, compound 4 was four times more active than chloroquine, while 3 showed very weak activity relative to natural quassinoids and positive controls. The in vitro sensitivity of the P. falciparum strain toward the compounds tested was similar and reproducible in assays in duplicate on separate occasions. Some quassinoid structural requirements, such as an α,β-unsaturated ketone in the A ring, an epoxymethylene bridge on the C ring and an ester functionality at C-15 are considered important for the antimalarial activity presented by quassinoids (Okano et al. 1995) and are in general present in the compounds under study. An ethnopharmacological study in French Guyana indicated that the P. sprucei root, stem and bark alcohol extract is used in local traditional medicine as a curative treatment for malaria in association with Geissospermum spp. (Apocynaceae) and Quassia amara (Simaroubaceae) or modern drugs (Vigneron et al. 2005). In a subsequent study, Bertani et al. (2005) demonstrated that P. sprucei water extract can inhibit hemozoin formation. In vitro assays demonstrated the antimalarial activity of this extract against the chloroquine-resistant P. falciparum strain W2. In the present study, we have demonstrated that both quassinoids 1 and 2 are isolated from infusions of P. sprucei in the laboratory procedure described and it is reasonable to assume that they are at least partially responsible for the antimalarial activity reported previously for P. sprucei water extracts by Vigneron et al. (2005) and Bertani et al. (2005). The erythrocytic membrane is a delicate structure that can be significantly altered by drug interactions (Aki & Yamamoto 1991). Several studies indicate that certain compounds isolated from plants such as polyphenols, glycosides, saponins and triterpenoids may cause changes in the membranes of red blood cells and subsequently produce haemolysis (Ng et al. 1986, Bader et al. 1996, Grinberg et al. 1997, Zhang et al. 1997). The mechanical stability of the erythrocyte membrane is a good indicator of in vitro insults in programs for cytotoxicity screening, since its structural dynamics favour interactions with drugs and changes that may be assessed by protein electrophoresis (Sharma & Sharma 2001). Erythrocytes also provide a simple model for the study of the protective or toxic effects of substances or situations associated with oxidative stress (Aparicio et al. 2005, Lexis et al. 2006, Muñoz-Castañeda et al. 2006). Our results suggest that the observed high cytotoxicity of quassinoids towards tumor cell lines is not due to oxidative stress or damage to the cell membrane, since none of the quassinoids or their derivatives were observed to lyse mouse erythrocytes at concentrations below 200 µg/mL. Natural quassinoids 1 and 2 display higher activities than their acetylated derivatives for all biological tests performed (except haemolytic activity, where as was stated above, no activity was observed for any of these compounds). This is evidence that the presence of a hydroxyl group at C-12 may be important for the cytotoxic, larvicidal and antimalarial activities observed in the present study. The enone group in ring A of substances 1 and 3 likely acts as a Michael acceptor for biological nucleophiles (Kupchan et al. 1975). This is strongly supported by the significant increase in biological activity associated with the presence of a free hydroxyl group at C-1 and/or C-3 in quassinoids (as 1 has a free hydroxyl group at C-1, while its inactive derivative 3 does not). This is presumably due to intramolecular hydrogen bonding between these hydroxyl groups and the enone oxygen atom, further activating the enone moiety towards nucleophilic attack (Guo et al. 2005). Additionally, diminished cytotoxic activity (28, 72, 95 and 122 times less against SF295, HL-60, MDA-MBA345 and HCT-8, respectively) of 4 (no free hydroxyl group at C-12) in comparison to 2 (free hydroxyl group at C-12) is evidence that the hydroxyl group at C-12 is crucial for the observed cytotoxicity (Table). Similar data are available for a number of quassinoids bearing A-D ring structures analogous to 1 and 2 and quantitative structure-activity analyses have attributed this cytotoxic activity to the presence of carbonyl and hydroxyl groups on the A ring, an epoxymethylene bridge between C-8-C-13 and a side-chain containing an ester (oxycarbonyl) group at C-15 (Okano et al. 1995). Quassinoids 1 and 2 from P. sprucei display important in vitro cytotoxic, antiplasmodial and larvicidal activities. The antiplasmodial activity described above, especially for 1, and anthelmintic activity previously described by us for 1 and 2 lend support to traditional medicinal practice and recent ethnobotanical and ethnopharmacological studies all of which indicate the efficacy of P. sprucei (Vigneron et al. 2005, Nunomura et al. 2006) In general, (di)acetylated semi-synthetic quassinoid derivatives 3 and 4 displayed less in vitro cytotoxic, antiplasmodial and larvicidal activity than the natural compounds from which they were derived. The effective larvicidal activity exhibited by quassinoids underscores the need for more specific tests for the possible effects on the peritrophic matrix structure, as antifeedants, and for effects of quassinoids on other larval stages. Tests for insecticide activity in A. aegypti imagoes (adults) and mosquitos of the Anopheles and other genera, studies of synergism and in vivo antitumor and antimalarial (including artemisinin resistant strains) activities and the mechanism of cytotoxic activity are underway and the results will be reported in due course. Additionally, the preparation of novel semi-synthetic quassinoid derivatives may provide compounds with more adequate physical and medicinal properties (stability, solubility, toxicity, specific action mechanisms) and more detailed information about the structure-activity relationships. Supplementary data ACKNOWLEDGEMENTS To the Centro de Biotecnologia da Amazônia for NMR spectra, to the the following financial agencies for schlorships received: VFAN (MCT/INPA/PCI), COP (CNPq), BCC (140095/2007-1), MRSM (CNPq 142115/06), ECCS (CAPES), RCNA (CNPq 142115/06-1), and to Walldelice H. Salgado and Michele Rodrigues from FMT-AM for technical support received. REFERENCES
Copyright 2009 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc09008t1.jpg] [oc09008f2.jpg] [oc09008f3.jpg] [oc09008f4.jpg] [oc09008f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}