 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 104, No. 1, February, 2009, pp. 93-99 Camallanus tridentatus (Drasche) (Nematoda: Camallanidae): new taxonomically important morphological data Cláudia Portes SantosI, +; František MoravecII ILaboratório
de Avaliação e Promoção da Saúde Ambiental,
Instituto Oswaldo Cruz-FIOCRUZ, Av. Brasil 4365, 21045-900 Rio de Janeiro, RJ,
Brasil Financial support: FAPERJ, PAPES V/FIOCRUZ, Grant Agency of the Czech Republic (524/06/0170) Received 18 September
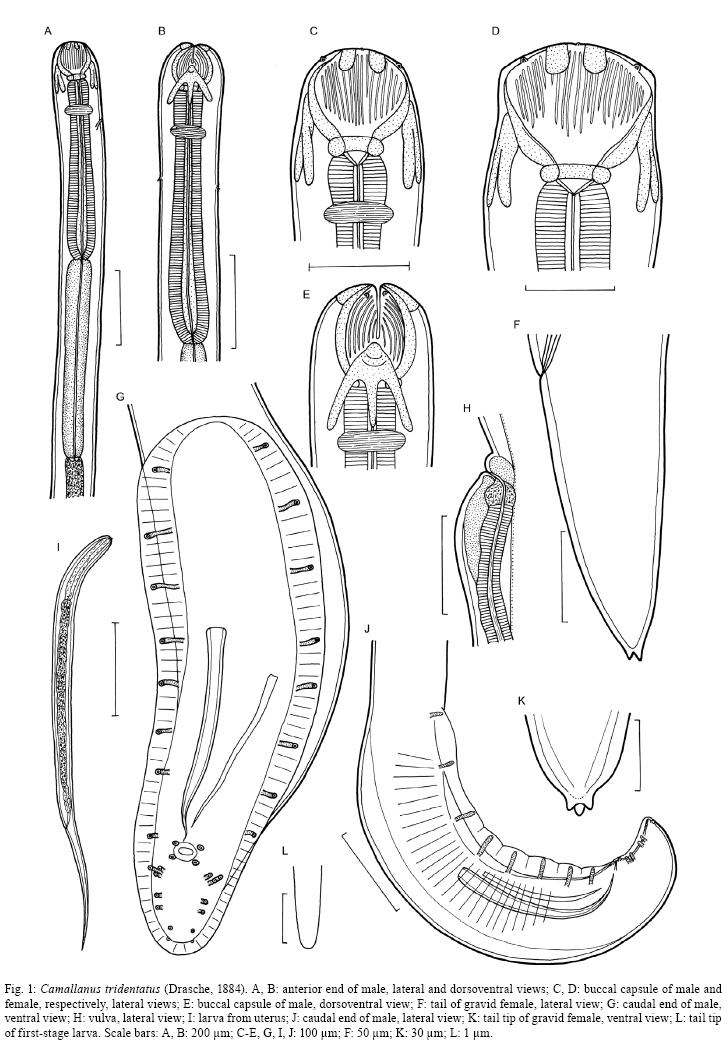
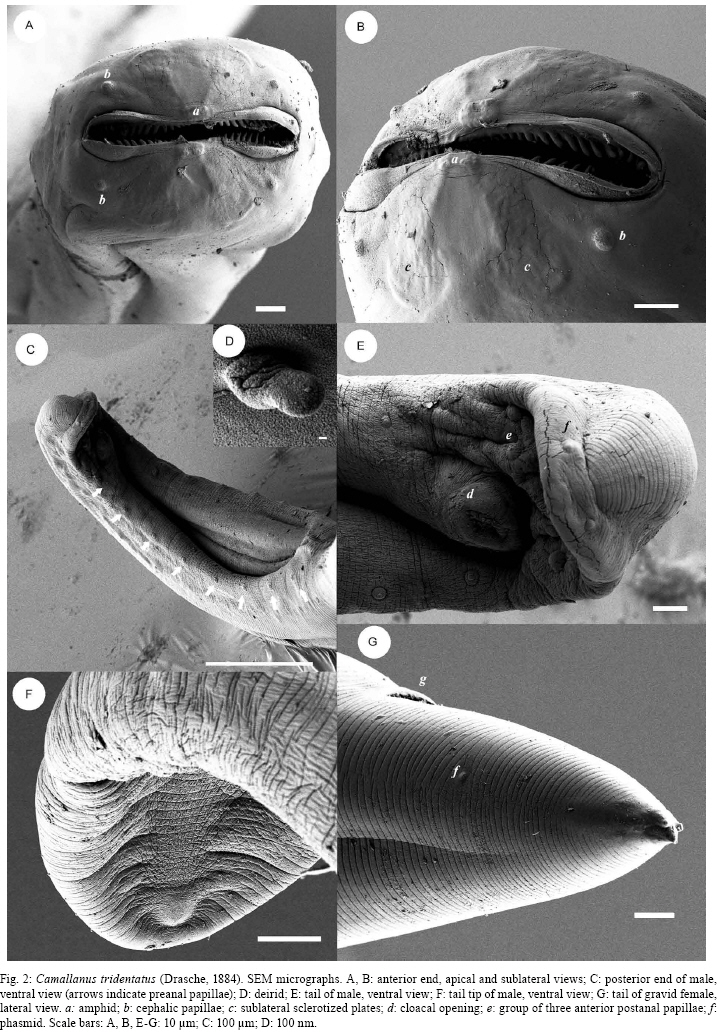
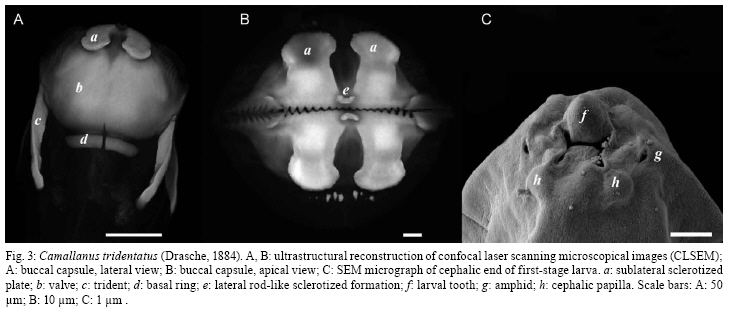
2008 Code Number: oc09014 ABSTRACT Camallanus tridentatus is redescribed on the basis of the examination of specimens obtained from the stomach, caeca and intestine of the naturally infected arapaima Arapaima gigas (Schinz) from the Mexiana Island, Amazon River Delta, Brazil. Data on the surface morphology of adults inferred from confocal laser scanning and scanning electron microscopical observations are also provided. The study revealed some taxonomically important, previously unreported morphological features in this species, such as the presence of the poorly sclerotized left spicule and deirids. C. tridentatus distinctly differs from other congeneric species parasitizing freshwater fishes in South America mainly in the structure of the buccal capsule and the female caudal end. C. maculatus Martins, Garcia, Piazza and Ghiraldelli is considered a junior synonymm of Camallanus cotti Fujita. Key words: Camallanus tridentatus - Nematoda - Arapaima gigas - Brazil During recent investigations into the helminth parasites of fish in the Mexiana Island, Amazonia, Brazil, specimens of Camallanus tridentatus (Drasche) were collected from the arapaima Arapaima gigas (Schinz). Drasche (1884) briefly described this species as Cucullanus tridentatus from a single female specimen found by him among Diesing's type material of Goezia spinulosa (Diesing) from A. gigas, originally collected by Natterer in Brazil. Later Baylis (1927) shortly described another, probably non-gravid female, of C. tridentatus, from a collection of helminth parasites of A. gigas sent to the then British Museum (Natural History) in London from Brazil. The very incomplete data on the females of C. tridentatus by these authors represented the only available knowledge of this parasite species for the following more than 70 years, until Ferraz and Thatcher (1990) published a redescription of C. tridentatus based on one male and six females collected in A. gigas from the Negro River, state of Amazonas (AM), Brazil. Based on the examination of freshly collected material, the species is redescribed herein and new data on the morphology of adults and first-stage larva are inferred from confocal laser and scanning electron microscopy observations. MATERIALS AND METHODS The fish were obtained from the Fazenda Santo Ambrosio area, Mexiana Island (Amazon River delta), state of Pará (PA) (Pirarucu Management Project, IBAMA 005-2007). A total of 30 specimens of arapaimas body length 6.5-175cm were examined for the presence of helminth parasites. The camallanid nematodes recovered were washed in physiological saline and then fixed in hot 4% formaldehyde solution. For light microscopy, the nematodes were cleared with glycerine. Drawings were made with the aid of a Zeiss drawing attachment. After examination, the specimens were stored in vials with 70% ethanol. Measurements are given in micrometres unless otherwise stated, with means in parentheses. For scanning electron microscopy (SEM), fixed specimens were postfixed in 1% osmium tetroxide in phosphate buffer, dehydrated through a graded acetone series, critical-point dried and sputter-coated with gold. The samples were examined using a JEOL ISM-7401F scanning electron microscope at an accelerating voltage of 4 kV or 8 kV. For confocal laser scanning microscopy studies, the specimens fixed in 4% paraformaldehyde were incubated overnight with faloidin conjugated to FITC 1:700 in PBS (Sigma) and mounted in semi-permanent slides in PBS with 2.5% de 1,4-diazabiciclo-(2,2,2)-octano-trietilenodiamina and 50% glycerol, pH 7.2. Studies were performed using an Olympus BX51 with Fluoview version 3.2. In addition to newly collected specimens from Mexiana Island, one stained gravid female of C. tridentatus (mounted as a slide), collected from A. gigas from the Negro River, AM, Brazil, by Ferraz and Thatcher (1990) and now deposited in the Helminthological Collection of the Institute of Parasitology, BC ASCR, in eské Budjovice (Cat. no. N-528) was examined. Moreover, the type specimens of Camallanus maculatus Martins, Garcia, Piazza and Ghiraldelli, 2007 (holotype, allotype and 14 paratypes), deposited in the Helminthological Collection of the Instituto Oswaldo Cruz, Rio de Janeiro (Cat. no. 35283), were re-examined. Fish names follow FishBase (Froese & Pauly 2008). Camallanus tridentatus (Drasche, 1884) Railliet, 1915 Syn. Cucullanus tridentatus Drasche, 1884 General diagnose: Camallanidae. Medium-sized nematodes with thick, finely transversely striated cuticle (Figs 2C, E, G) and large, orange-brown capsule typical of genus, i.e., consisting of two sclerotized lateral valves internally supported by longitudinal ridges and each bearing two short sclerotized plates at anterior margin, short basal ring, and well-developed dorsal and ventral tridents (Figs 1A-E, 3A, B). Body of larger females reddish (brownish after fixation), with distinct dark-brown intestine; body of small specimens whitish. Mouth aperture slit-shaped, dorsoventrally elongated, surrounded by four large submedian cephalic papillae and pair of small lateral amphids (Fig. 2A, B). Valves of buccal capsule roughly pentagonal in lateral view, each internally bearing 19-26 smooth longitudinal ridges not reaching posteriorly to basal ring; some ridges (usually 3-5) may be much shorter than others; anterior ends of ridges protruding into mouth as small teeth, resembling a jaw (Figs 2A, B, 3B). Sublateral superficial sclerotized plates extending posteriorly to about fourth of length of valves (Figs 1C, D, 3A); their posterior ends distinctly expanded (Fig. 3B). Additional small, superficial transverse rod-like sclerotized formation present at top of each valve (between bases of two sublateral plates), forming a kind of support of amphid (Fig. 3B). Bottom of capsule formed by well-developed, sclerotized basal ring. Tridents large, surpassing posterior border of buccal capsule. Distinct oesophageal cup with colourless sclerotized walls present between buccal capsule and muscular oesophagus (Fig. 1C, D). Muscular oesophagus slightly shorter than glandular oesophagus; posterior part of former somewhat expanded (Fig. 1A, B). Excretory pore situated short distance posterior to level of nerve ring. Deirids small, simple, situated some distance posterior to level of nerve ring (Figs 1B, 2D). Male (9 specimens): Length of body 7.30-8.38 (7.88) mm, maximum width 163-204 (154). Buccal capsule including basal ring 80-120 (95) long, maximum width 87-132 (96); basal ring 10-18 (14)long, 45-75 (55) wide; length of tridents 77-90 (81). Each valve of capsule strengthened internally by 19-24 (21) (longitudinal ridges, 3-5 of them being markedly shorter than others. Oesophageal cup 9-15 (11) long, 18-24 (21) wide. Muscular oesophagus 488-585 (522) long, 84-96 wide (90); glandular oesophagus 620-789 (660) long, 60-69 (64) wide; length ratio of both parts of oesophagus 1:1.2-1.3. Length of buccal capsule and oesophagus forms 16-19% of total body length. Nerve ring and deirids 175-210 (185) and 312-315 (313), respectively, from anterior extremity; excretory pore not located. Caudal end of body conspicuously broad, with vesiculate caudal alae 475-580 (525) long supported by pedunculate papillae; caudal alae joining anteriorly to form a kind of ventral pseudosucker (Figs 1G, J, 2C). Caudal papillae: 8-9 pairs of preanal and 6 pairs of postanal pedunculate papillae present. Preanal papillae: 8-9 pairs of subventral, equally spaced papillae. Postanal papillae: six pairs of subventral papillae; first three pairs close to each other, following two pairs equally spaced; last pair near to tail tip. Cloacal opening surrounded by two pairs (1 precloacal and 1 postcloacal) of additional, small sesille papillae (Fig. 1G, J). Pair of small papilla-like phasmids situated dorsally near to tail tip (Fig. 2E). Spicules unequal, simple, with sharply pointed distal ends (Fig. 1G, J). Large (right) spicule heavily sclerotized, orange-coloured, 212-276 (225) long; small (left) spicule poorly sclerotized, colourless, 90-105 (98) long. Tail conical, 85-135 (106) long, with rounded tip without any outgrowths (Fig. 2F). Female (11 gravid specimens): Body length 12.50-14.77 (14.28) mm, maximum width 280-408 (328). Buccal capsule including basal ring 132-171 (150) long, maximum width 125-192 (156); basal ring 15-24 (21) long, 70-90 (78) wide; length of tridents 115-140 (130). Each valve of capsule with 24-26 (25) longitudinal ridges. Oesophageal cup 15 (15) long, 24-30 (27) wide. Muscular oesophagus 700-870 (850) long, 123-138 (128) wide; glandular oesophagus 860-993 (926) long, 75-96 (84) wide; length ratio of both parts of oesophagus 1:1.0-1.2. Length of buccal capsule and oesophagus forms 12-14% of total body length. Nerve ring, excretory pore and deirids 242-279 (267), 299-381 (340) and 598, respectively, from anterior extremity. Vulva postequatorial, 7.17-8.70 (7.90) mm from anterior extremity, at 55-59% of body length, with somewhat elevated vulval lips. Vagina muscular, 300-450 (353) long, directed posteriorly from vulva (Fig. 1H). Uterus filled with numerous larvae. Larvae (n = 5) 483-501(492) long, maximum width 18 (18); cephalic end blunt, bearing four submedian cephalic papillae, pair of lateral amphids and dorsal larval tooth; tail slender, 156-168 (162) long (32-34% of body length), with rounded tip without any outgrowths (Figs 1L, 3C); oesophagus 75-90 long (representing 15-19% of larval body length). Female tail conical, 140-245 (192) long, representing 1.1-1.7% of body length; pair of small lateral phasmids situated at approximately its middle (Fig. 2G). Tail tip provided with 3 minute caudal projections; dorsal projection longer (6) than both subventral projections (3) (Figs 1F, K, 2G). Host - Arapaima, A. gigas (Arapaimidae, Osteoglossiformes), local name "pirarucu", body length 70-175 cm. Sites of infection - Stomach, pyloric caeca and anterior part of intestine. Locality - Natural canals at Fazenda Santo Ambrosio (00º05'30"S, 49º34'50"), Mexiana Island (Amazon River delta), PA, Brazil. Prevalence and intensity - 33% (10 fish infected/30 fish examined); 1-170 nematodes per fish. Fish smaller than 70cm had no camallanids. Depository of voucher specimens - Instituto Oswaldo Cruz-Fiocruz, Rio de Janeiro, Brasil (CHIOC 35611). DISCUSSION C. tridentatus was described by Drasche (1884) and briefly redescribed by Baylis (1927) from A. gigas from Brazil. Later, Ferraz and Thatcher (1990) also redescribed this nematode from the same host species. Several authors reported C. tridentatus from the perciform fishes Percichthys trucha (Valenciennes) and Percichthys colhuapiensis MacDonagh and from salmonids from Argentina but, apparently, another species, Camallanus corderoi Torres, Teuber and Miranda was mistaken for C. tridentatus (Ortubay et al. 1994, Moravec 1998). The present study, for the first time based on numerous specimens of C. tridentatus and using, in addition to light microscopy, also the methods of scanning electron and confocal laser scanning microscopy, has revealed some taxonomically important, previously not reported morphological features in this species. Although Ferraz and Thatcher (1990) observed only one spicule (205 long) in the male of C. tridentatus, the present study shows that there are two spicules in this species. This confirms the opinion of Moravec et al. (2006) that the presence of two spicules is a generic feature in Camallanus Railliet. Generally, in contrast to the well-sclerotized right spicule, the left spicule in camallanids (e.g., Camallanus, Procamallanus Baylis) is only weakly sclerotized; in some species it is so poorly sclerotized that it is hardly visible and, in such cases, it may appear to be completely absent. Re-examinations of some species reported to possess only one spicule showed that also the left spicule was present (De & Moravec 1980, Moravec et al. 2003, 2006). In this study, the structure of the buccal capsule of C. tridentatus has been found to be typical of Camallanus. Although Drasche (1884) and Baylis (1927) reported the number of longitudinal ridges per valve in C. tridentatus females to be about 30, the present data show somewhat lower numbers (19-26), as already indicated by Ferraz and Thatcher (1990). SEM and confocal microscopy examinations have shown that the anterior ends of these ridges form small teeth protruding into the mouth cavity, being arranged as jaws; the sublateral plates have been found to grow out of the anterior margins of the valves and small lateral, sclerotized transverse rod-like formations have been observed, for the first time in a Camallanus spp., at the tops of valves. Confocal laser scanning electron microscopy proved to be very useful for the examination of the buccal capsule surface in C. tridentatus and it would be desirable to use this method for other Camallanus spp. for comparative purposes; recently, Rigby et al. (2008) achieved similar results when studying the surface of the buccal capsule in two newly described Camallanus spp. with the use of a digestive method (acid-pepsin) to remove the tissues surrounding the capsule and the subsequent examination of the capsule by SEM. Ferraz and Thatcher (1990) reported the presence of 14 pairs (8 preanal, 2 adanal and 4 postanal) of caudal papillae in the male of C. tridentatus, but they did not observe them in a ventral view. In the present material, the males had a larger number of pairs of papillae (16-17), eight (9 in one male) preanal, two circumanal and six postanal; this arrangement of papillae, especially the presence of two small sessile circumanal papillae and the group of the first three pairs of pedunculate postanal papillae, is probably characteristic of the most species of Camallanus (Moravec et al. 2004b, 2006, Moravec & Justine 2006). The caudal tip of the female of C. tridentatus was described by Ferraz and Thatcher (1990) as bifid, but in fact it bears three small conical processes, as visible from SEM micrographs, which are also present at the female tail tip of some other Camallanus spp. The presence of amphids, deirids and the body size of the first-stage larvae from the uterus are described in C. tridentatus for the first time in this paper. In contrast to Camallanus lacustris (Zoega) and Camallanus cotti Fujita, in which the first stage-larva possesses outgrowths on the tail tip (Moravec & Justine 2006), such outgrowths are absent from the first-stage larva of C. tridentatus (Fig. 1L); such caudal outgrowths were neither found in the first-stage larva of Camallanus oxycephalus Ward and Magath studied by SEM (Kelly et al. 1989). To date, only three valid species of Camallanus, Camallanus acaudatus Ferraz and Thatcher, C. corderoi and C. tridentatus are known as parasites of native freshwater fishes in South America (Moravec 1998). Whereas C. corderoi, a parasite mainly of P. trucha in Chile and Argentina, has the buccal capsule with dentigerous ridges (subgenus Zeylanema Yeh), both other species, parasites of osteoglossiform fishes in the Amazon River Basin, have smooth ridges in the capsule (nominotypical subgenus Camallanus); the latter differ from each other in conspicuously different lengths and shapes of the female tail. Recently, Martins et al. (2007) described a new species, C. maculatus Martins, Garcia, Piazza and Ghiraldelli from the cultured ornamental fish Xiphophorus maculatus (Günther) in Brazil, but this is evidently a synonym of C. cotti, an Asian species now introduced into many countries of different continents, where it parasitizes many species of cultured and native fish species (Moravec & Justine 2006, Wu et al. 2007). Moravec and Justine (2006) suggested this synonymy, but because the paper describing C. maculatus was not yet published at that time, they could not do it formally. After a re-examination of the type specimens (male holotype, female allotype and 14 paratypes) of C. maculatus, it is possible to confirm the synonymy of this species with C. cotti. C. cotti differs from all the above mentioned South American species of Camallanus, principally in possessing an unusually long female tail without caudal processes. The pathogenicity of camallanid nematodes is not sufficiently known, but it appears to be manifested mainly by the loss of host's blood, on which the parasites feed and in the mechanical damage to the intestinal mucosa (Moravec 1994). The pathogenicity of C. tridentatus has not yet been studied; however, high infections by some species of Camallanus and Procamallanus are known to be responsible for a complete destruction of the mucosa of the host's digestive tract, which may lead to the death of the fish host (Petter et al. 1974, Stumpp 1975, Campana-Rouget et al. 1976, Machida & Taki 1985, Moravec et al. 2004b). Although the prevalence of C. tridentatus in arapaimas of the Mexiana Island was high (33%), the intensity varied from 1-170 nematodes per fish; taking into account that only large fish (70-170 cm long) were infected with this parasite, it can be assumed that C. tridentatus did not seriously affected the health condition of the infected fish. However, it may be different under certain conditions of a fish culture, where also small fish may be heavily infected, as it was recorded, for example, for Procamallanus guttatusi (Andrade-Salas, Pineda-López and García-Magaña) infecting the marine fish Siganus guttatus (Bloch) cultured in farms in the Philippines (Machida & Taki 1985, Moravec et al. 2004a). The presence of C. tridentatus only in large specimens of A. gigas suggests that paratenic hosts (probably various small fishes) are involved in the life cycle of this nematode, representing the main source of infection for arapaimas. ACKNOWLEDGEMENTS To Dr. Luiz Rabelo Neto, from Grupo Reicon, and Dr. Rossana Venturieri, for the facilities and support provided in field work, to Dr. Luis Muniz, Helminthological Collection of the IOC-Fiocruz, kindly provided the specimens for study, to Dr. Marcelo Pelajo and Pedro Paulo de Abreu Manso, IOC-Fiocruz, for providing the Laser Scanning Electron Microscopic facilities, and to the staff of the Laboratory of Electron Microscopy, Biology Centre, ASCR, eské Budjovice, for technical help. REFERENCES
Copyright 2009 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc09014f1.jpg] [oc09014f3.jpg] [oc09014f2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}