 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 104, No. 1, February, 2009, pp. 124-126 SHORT COMMUNICATIONS Embryonic development of Aedes aegypti (Diptera: Culicidae): influence of different constant temperatures Luana Cristina Farnesi; Ademir Jesus Martins; Denise Valle; Gustavo Lazzaro Rezende+ Laboratório
de Fisiologia e Controle de Artrópodes Vetores, Instituto Oswaldo Cruz-
Fiocruz, Rio de Janeiro, RJ, Brasil, Laboratório de Entomologia, Instituto
de Biologia do Exército, Rio de Janeiro, RJ, Brasil Financial support: CNPq, FAPERJ, FIOCRUZ Received 14 October
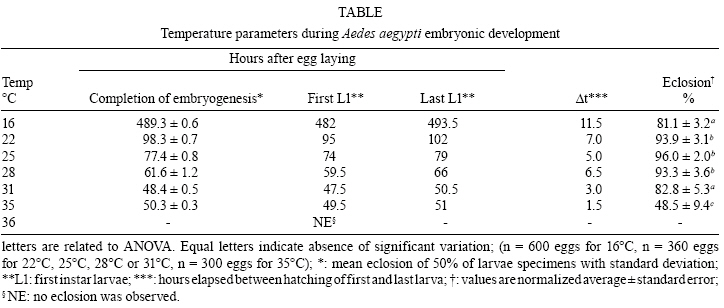
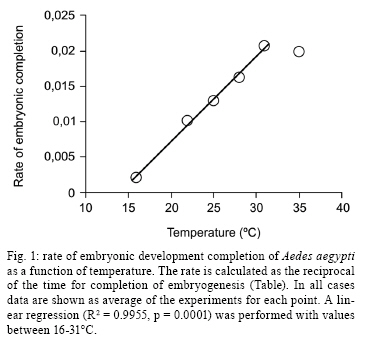
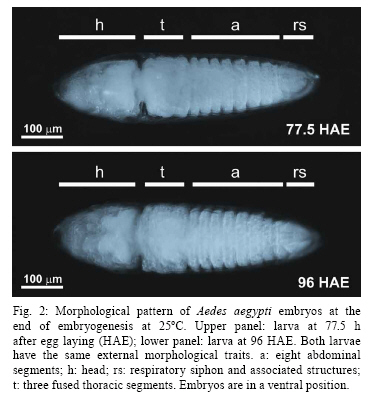
2008 Code Number: oc09020 ABSTRACT Despite its vector importance little attention is given to Aedes aegypti embryonic development. In this study, temperature influence on time course of Ae. aegypti larvae hatching and egg viability were evaluated. The dormancy state at the end of embryogenesis could be interrupted with a proper stimulus. Temperatures tested ranged between 12-36°C; the maximum temperature limit is 35°C and the minimum one is below 12°C. Egg viability between 16-31°C was above 80%. The definition of physiological embryonic parameters at this temperature range corroborates Ae. aegypti presence on tropical and subtropical world regions. Keywords: Aedes aegypti - embryogenesis - temperature influence Dengue and yellow fever are considered two of the most important viral diseases transmitted by arthropods (Gould & Solomon 2008). The mosquito Aedes aegypti, vector of urban yellow fever and also the main dengue vector, is widely distributed worldwide, being usually found between latitudes 35° N and 35° S (OPAS, 1995). Ae. aegypti embryos tend to enter dormancy at the end of embryogenesis, being able to keep its viability inside the egg for six months or more, according to the environmental relative humidity (Christophers 1960, Kliewer 1961, Clements 1992). When the surroundings become favorable to larval development, e.g. flooding of larvae habitat, dormancy is interrupted and the larva hatches (Christophers 1960, Clements 1992). Little attention has been given to the influence of temperature on Ae. aegypti embryonic development, when compared to other mosquito species (Rosay 1959, Christophers 1960, Trpiš et al. 1973, Clements 1992, Gillooly & Dodson 2000). The precise time Ae. aegypti takes to complete its embryogenesis at distinct temperatures is not known (Christophers 1960, Consoli & Lourenço-de-Oliveira 1994). In the present work the influence of distinct constant temperatures on Ae. aegypti embryogenesis, egg viability and the time course of larval hatching were analyzed. To induce Ae. aegypti larval eclosion from eggs previously stored in our laboratory for one month, preliminary tests were performed utilizing conditions described as effective stimuli in the literature (Gander 1951, Barbosa & Peters 1969, Novak & Shoroyer 1978). In all cases mosquitoes from the Rockefeller strain were used. The conditions used were: (i) dechlorinated water; (ii) deaerated dechlorinated water; (iii) 0.3 mM ascorbic acid in dechlorinated water; (iv) 0.3% (w/v) yeast extract (SIGMA # Y1625) solution in dechlorinated water (YES); (v) 0.3% (w/v) yeast extract solution in deaerated water. All solutions above were freshly made. The best stimulus for Ae. aegypti larval hatching, in our laboratory conditions, is the 0.3% (w/v) YES, the rate of egg hatching being 80% on average (data not shown). We observed that the embryonic dormancy state of Ae. aegypti is facultative, as expected for a multivoltine species (Forattini 2002), and could be bypassed by adding YES at the end of embryogenesis. Preliminary experiments have been performed in order to time this event, by adding YES at distinct hours after egg laying. Synchronous egg laying was performed during 20 min, as recently described (Rezende et al. 2008). After this period, eggs were kept at the distinct temperatures employed in the experiments. The precise moment of hatching of the first larvae was then observed for each temperature (see Table for the values). For all further experiments, YES was added 2 h before this moment. In this way, it was possible to monitor Ae. aegypti eclosion immediately after completion of embryonic development. For each experiment at a constant temperature, triplicates from the same egg laying were utilized. These eggs were kept on moist Whatman # 1 paper until YES was added. From this moment on, larvae hatching was observed every 30 min, until no more hatching occurred. Three experiments were performed for each tested temperature, on different days and with different egg batches, except at 16ºC and 35°C when two experiments were performed. Relevant parameters of embryogenesis at distinct temperatures are shown on Table. Eclosion rates above 90% were observed between 22-28°C. These values dropped to 80% at 16ºC and 31°C and at 35°C the eclosion rate was approximately 50%. No hatching was observed above 36°C, although viability tests at 26°C indicate normal eclosion of eggs from the same egg batches. Therefore, 35°C was defined as the maximum temperature supported for embryonic development of Ae. aegypti. The velocity of the completion of embryonic development is directly proportional to the temperature, except at 35°C, where completion is slower than at 31°C (Table, Fig. 1). Our hypothesis is that this delay is due to a greater physiological stress during embryogenesis, caused by the high temperature at which Ae. aegypti embryos were exposed. The same phenomenon is observed during larval development of this species, which takes longer at 33°C than at 31°C (Gilpin & McClelland 1979). After about 960 h (40 days) egg hatching was observed at 12°C. Clarification of eggs (n = 300) kept at 12°C for 13 days (approximately 312 h), indicated embryos were still in early stages of development (data not shown). Christophers (1960) observed that 7°C completely inhibits Ae. aegypti embryogenesis. Therefore, while the minimum temperature supported by this species during embryonic development has not been determined, it can be stated that it falls between 7-12°C. In any case, the definition of Ae. aegypti physiological embryonic parameters for temperatures between 12- 35°C is in accordance with the distribution of urban yellow fever and dengue in the tropical and subtropical regions of the globe (Guzmán & Kourí 2002, Barrett & Higgs 2007). Using the hatching stimuli methodology with YES, we found Ae. aegypti embryonic development at 25°C is completed at 77.5 h. However, based on morphological parameters, Raminani and Cupp (1978) previously reported Ae. aegypti embryogenesis takes 96 h at this temperature. With Trpiš clarification solution (Trpiš 1970, Rezende et al. 2008) we analyzed whether there were apparent morphological differences between embryos 77.5 h and 96 h old. In both cases embryos displayed the same external morphological characters, i.e., separated head, three fused thoracic segments, eight abdominal segments and respiratory siphon with associated structures (Christophers 1960) (Fig. 2). Additionally, embryos hatched at 77.5 h were followed up to the end of pupae stage, exhibiting similar viability rate to embryos left to hatch at 96 h. 70.5% and 73.9% of embryos hatched at 77.5 h and 96 h developed up to the pupa stage, respectively (n = 120 eggs for each condition). Development of the hatching spine, the larval structure that assists eggshell opening in the course of hatching, is accomplished 75 h after egg laying at 25°C (Raminani & Cupp 1978). This observation led us to hypothesize that presence of a fully developed hatching spine could be an indication that larvae are ready to hatch, which eventually happens when the YES is added. In warm and humid countries, such as Brazil, dengue is still a serious public health problem, where environmental conditions favor the development and proliferation of its vector (Lourenço-de-Oliveira 2008). There is an influence of temperature on development and reproduction of Ae. aegypti populations in the wild. Field studies correlate a temperature effect over mosquito abundance (Lourenço-de-Oliveira 2008, NA Honório, personal communication) or even with the absence of oviposition activity and immature forms during the winter (Vezzani & Carbajo 2008). Moreover, knowledge of the effect of temperature on the embryonic development of culicids may serve as a basis for other studies such as the possibility of systematic evaluation of gene expression and physiological changes during embryogenesis (Rezende et al. 2008). The precise identification of embryonic kinetics can also assist the development of new alternatives to the control of disease vectors, based on information obtained from embryological studies. ACKNOWLEDGEMENTS To Jutta Gerlinde Birgitt Linss, for translating the Gander paper, and Rafaela Vieira Bruno and the reviewers, for critical reading and suggestions on the manuscript. REFERENCES
Copyright 2009 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc09020t1.jpg] [oc09020f1.jpg] [oc09020f2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}