 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 104, No. 2, March, 2009, pp. 149-154 Brazilian contribution for a better knowledge on the biology of Toxoplasma gondii Wanderley de SouzaI, +; Renato A DaMattaII; Márcia AttiasIII IInstituto
Nacional de Metrologia, Normalização e Qualidade Industrial, Rio
de Janeiro, RJ, Brasil Received 10 October
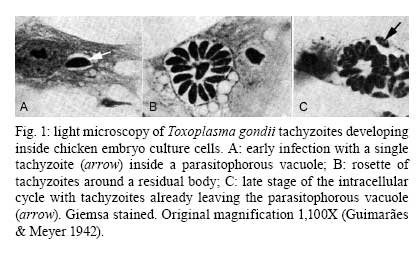
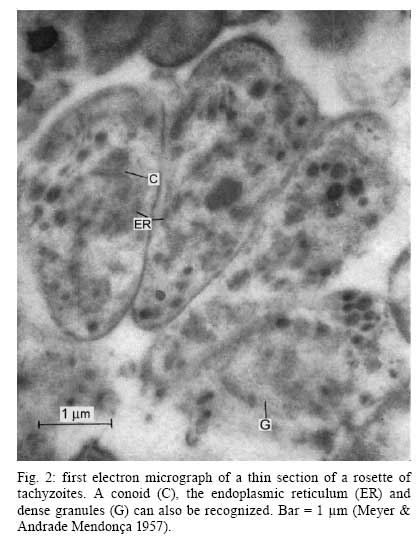
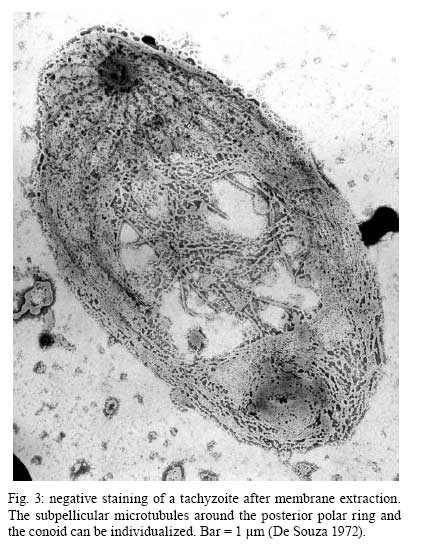
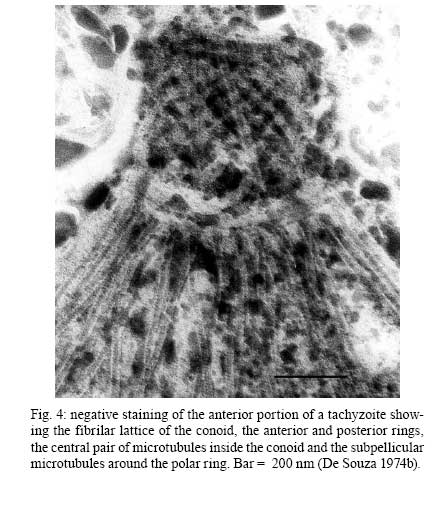
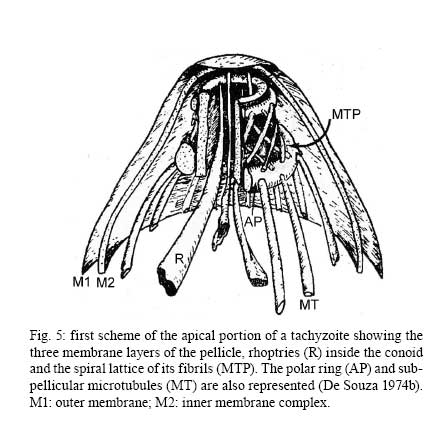
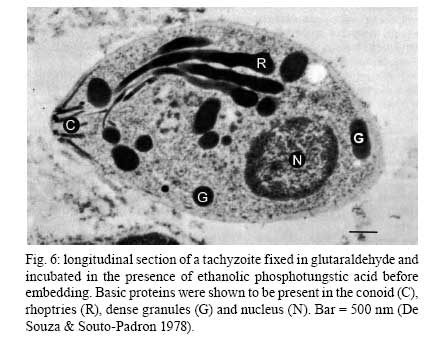
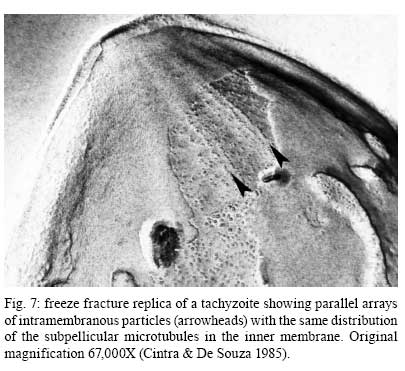
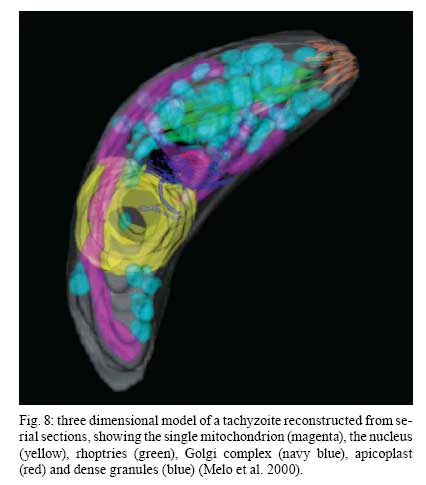
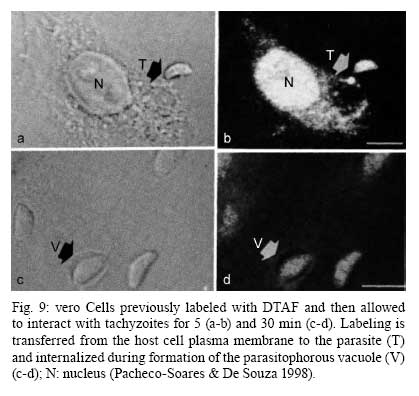
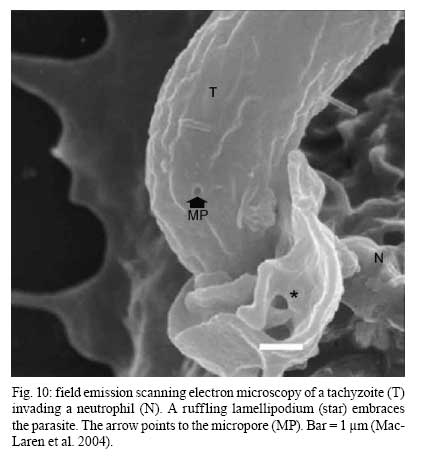
2008 Financial support: CNPq, FINEP, FAPERJ Code Number: oc09027 ABSTRACT Historically, scientists in Brazil has significantly contributed to the biology, cultivation and structural organization of the pathogenic protozoan Toxoplasma gondii and its interaction with host cells, starting with the description of the protozoan by Splendore in 1908. The intracellular and extracellular corpuscoli observed in rabbits, corresponded to what we now as tachyzoites. Later on, a pioneering method to grow T. gondii in tissue cultures was developed by Guimarães and Meyer, 1942. They also observed for the first time T. gondii by transmission electron microscopy and made the initial description of the cytoskeleton of T. gondii by observing negatively stained cells. In the 1980's, the relation of the cytoskeleton with the sub-pellicular microtubules was reveled by freeze-fracture. More recently, several Brazilian groups have analyzed in detail basic aspects of the early interaction of the protozoan with the host cell, such as the role of protein phosphorylation, transfer of host cell surface components to the protozoan and genesis and organization of the parasitophorous vacuole. Tachyzoites strategically inhibit nitric oxide production during active invasion of activated macrophages. In vitro studies on the sexual cycle of T. gondii using primary cultures of cat enterocytes and the egress from host cells are being carried out. Perspectives are that the contribution of Brazilian science to the knowledge on T. gondii biology will continue to flourish in years to come. Key words:Toxoplasma gondii - Brazilian contribution - ultrastructure - host cell interaction The end of the XIX and the beginning of the XX century (30 years period) were a landmark for the biomedical sciences when several new important parasitic protist species, many identified as causal agents of important human diseases, were discovered. As a matter of fact, in 1880, Laveran described the genus Plasmodium and later, in 1903, the same Laveran, described Leishmania donovani. In the sequence, in 1907, Forde described Trypanosoma gambiense and, in 1908, Toxoplasma gondii was described. In 1909, Chagas described Trypanosoma cruzi and Pneumocystis carinii. In 1911, Viana described Leishmania braziliensis. For the Brazilian science, the first 10 years of the XX century were remarkable since Brazilian scientists were involved in the description of T. gondii, P. carinii, T. cruzi and L. braziliensis. As far as T. gondii is concerned, in 1909 Splendore, at that time working as director of the Bacteriological Laboratory of Hospital Português, in São Paulo, Brazil, had the opportunity to examine rabbits presenting an acute infectious process which lead to death. Pathological examination showed the presence of speciali corpuscoli, well stained with Giemsa, distributed in several tissues. The size of the corpuscoli was 5-8 μm long and 2.5-4 μm wide. Splendore sent the material to the great protozoologist Prowazek, at that time working in Rio de Janeiro. Both agreed that the corpuscoli belonged to a new group of protozoa. The communication was published in the Revista da Sociedade Scientifica de São Paulo, n. 10-12, 1908. In the same year, Nicolle and Manceau found the same protozoan in the rodent Ctenodactylus gondii captured in Tunis. In 1909, both Splendore (1909) and Nicole and Manceau (1909) published separately two other papers supporting the initial description. The new genus and the new species were designated by Nicolle and Manceau as T. gondii and Toxoplasma cuniculi. In 1914, Arantes made an important survey on the available data on T. gondii. His work was presented as a Doctoral Thesis to the Rio de Janeiro School of Medicine. Another important Brazilian contribution was the initial description, in 1927, by Magarinos Torres, a brilliant pathologist working at the Instituto Oswaldo Cruz, of congenital toxoplasmosis. Cultivation - A second major historical contribution from the Brazilian science was published in 1942, by Guimarães and Meyer, who set up a methodology for the cultivation of T. gondii in tissue culture. At that time, FN Guimarães and Hertha Meyer (1942), who initially worked with Fisher in Berlin and Levi in Turin, set up a tissue culture laboratory at the Instituto de Biofísica at the Federal University of Rio de Janeiro and made the first reproducible studies on the cultivation of T. gondii (Fig. 1A, B, C), T. cruzi and Plasmodium gallinaceum, reporting her observations in papers published in the 40's. More recently, Barbosa et al. (2005), working at the Instituto Oswaldo Cruz, in Rio de Janeiro, made the first attempts, using primary cultures of intestinal cells from cat embryos, to reproduce in vitro the sexual cycle of the protozoan. The fine structure - Meyer and Mendonça (1955, 1957) were among the first to use transmission electron microscope to analyze the fine structure of T. gondii, T. cruzi and P. gallinaceum. Indeed, the work published in 1957 (Meyer & Mendonça 1957) was the first to describe long, homogeneous dark stained inclusions, which clearly correspond to the rhoptries and dense granules (Fig. 2), now recognized as characteristic of the Apicomplexa phylum. One of us (WS) had the opportunity to join the group in 1969 and initiated a series of studies. The use of the negative staining technique allowed the visualization of the whole cytoskeleton composed of sub-pellicular microtubules (Fig. 3) and the sub-structure of the conoid (Fig. 4). These observations lead to the suggestion that this structure is made of rings and microtubules and change its shape and length (Fig. 4a), resulting in the proposition of a three-dimensional model for its structure (Fig. 5) (De Souza 1972, 1974a). After the initial morphological descriptions by transmission electron microscopy in the 1970's and 1980's several papers produced by Brazilian groups were focused on the application of new techniques to the study of T. gondii tachyzoites. Of particular interest, even today, is the report of the selective staining of secretory organelles (rhoptries, micronemes and dense granules) as well as the conoid and the subpelicular microtubules by ethanolic phosphotungstic acid, specific for the detection of basic proteins (Fig. 6) (De Souza & Souto-Padron 1978). This preparation method is particularly valuable to produce semithin sections for reconstruction of these structures by electrontomography. Freeze fracture of tachyzoites (Cintra & De Souza 1985) also showed intramembranous particles distributed in parallel arrays with the same spacing and periodicity of the subpelicular microtubules (Fig. 7), establishing an ultrastructural relationship between them and the inner membrane complex, as proposed in the models for gliding motility (reviewed in Baum et al. 2006). These observations were reinforced when protein bridges between the inner membrane complex and the subpellicular microtubules were also found in detergent extracted tachyzoites fixed in the presence of tannic acid. Furthermore, freeze fracture of infected cells treated with filipin showed the presence of cholesterol, an important component that is obtained from host cells, in the plasma, but not in the inner membrane complex of tachyzoites (Cintra & De Souza 1985). More recently, the ultrastructure and cell biology of the tachyzoite was reexamined and the activity of the single mitochondrion (Fig. 8) of intra and extra cellular tachyzoites was show with the use of the fluorescent compound CMXRos and observation by laser scanning confocal microscopy and computer aided 3-D reconstruction from serial sections, two cutting edge approaches at that time (Melo et al. 2000). A multiple technical approach, including light and transmission electron microscopy, freeze fracture, field emission scanning electron microscopy and 3-D reconstruction was also employed by Monteiro et al. (2001) in the analysis of the conoid of tachyzoites in the inserted and extruded states. Interaction with host cell - This is an area where several Brazilian groups have worked in the last years. Employing a chronological order, and considering primarily an ultrastructural approach, after the works by Silva et al. (1982), Meirelles et al. (1983) and Meirelles and De Souza (1985) on basic aspects of the invasion and survival of T. gondii in primary cell lines; the effect of various digestive enzymes and the cytochemical localization of plasma membrane enzyme markers during interiorization of tachyzoites of T. gondii by macrophages have been analysed by De Carvalho and De Souza (1989). The role of anionic sites, sugar residues and protein phosphorilation was covered in the works by De Carvalho and De Souza (1990), De Carvalho et al. (1991), Stumbo et al. (2002) and Ferreira et al. (2003). One important contribution, whose results have not been completely explored up to now, was made in experiments where the surface of host cells were labeled before tachyzoites invasion. After binding of the parasite to the host cell, the labeled lipids, proteins and glycoconjugates were transferred to the protozoan surface and subsequently found in the parasitophorous vacuole (Pacheco Soares & De Souza 2000) (Fig. 9). The redistribution of mitochondria and endoplasmic reticulum in host cells infected with T. gondii has also been shown by de Melo et al. (1992) and, more recently, by Magno et al. (2005). The host cell cytoskeleton was also shown to take part in the establishment of the parasitophorous vacuole by Andrade et al. (2001). More recently, the intranuclear localization of tachyzoites was also reported by Barbosa et al. (2005). This same group has reported that infection of primary cultures of skeletal muscle cells of mice with bradyzoites is a model for T. gondii cystogenesis (Guimarães et al. 2008) Field emission scanning electron microscopy has brought new insights and amazing views of the process of invasion of neutrophils (Fig. 10) (MacLaren et al. 2004) and the intravacuolar behavior of tachyzoites in non phagocytic cells (Magno et al. 2005) with special relevance on the organization and putative structural role of the intravacuolar network of tubular membranes. Interaction with macrophages - One of the legacies of Hertha Mayer, as previously mentioned, was cell culture. At that time, chicken embryos were easily obtained and were used, resulting in the hang drop cell culture technique. Another host cell model was chicken macrophages derived form blood monocytes. This cell type was used later on by one of us (WS) to show that chicken macrophages could destroy trypomastigotes of T. cruzi and promastigotes of Leishmania, but not tachyzoites of T. gondii (Meirelles & De Souza 1985). This experimental model was used also in an attempt to find out if nitric oxide (NO) was involved in this killing. It was shown that NO was not related to the killing of T. cruzi, actually it was the respiratory-burst activity that was responsible for the microbicidal action of these macrophages to T. cruzi (DaMatta et al. 2000). As expected, the survival of T. gondii in these macrophages was due to its evasion mechanism on the oxygen radicals burst (Murray & Cohn 1979, DaMatta et al. 2000). As a NO production control, mice macrophages activated with interferon-gamma and lipopolysaccharide were used and, in an unexpected way, it was reported that infection by T. gondii reduced NO production (DaMatta et al. 2000). Following this line of evidences, it was shown that a reduction of NO production correlated with higher infection rates in peritoneal macrophages of mice (Seabra et al. 2002) and also in chicken macrophages cell lines (Guillermo & DaMatta 2004). In both systems, inducible NO synthase (iNOS) disappeared only in infected macrophages and only live parasites were capable of inhibiting NO production. This inhibition occurred after infection independently if macrophages were activated before of after infection (Seabra et al. 2002, Guillermo & DaMatta 2004). The molecular mechanism behind this phenomenon was further investigated using the mice model. T. gondii infection induced a TGF-beta response, which lead to the disappearance of iNOS, explaining the inhibition of NO production (Seabra et al. 2004b). The secretion of TGF-beta was related to exposure of phosphatidylserine by tachyzoites (Seabra et al. 2004b). It was also shown that NFκB get out of the nucleus of activated macrophages and actin was depolymerized upon infection (Seabra et al. 2004b). However, the lack of lysosome fusion to the parasitophorous vacuoles was not related to the TGF-beta response, indicating that the evasion mechanism of T. gondii is multi-faceted. Unexpectedly, infection of activated macrophages by T. gondii did not increase the synthesis of polyamines as seen by monitoring ornithine descarboxilase activity after infection (Seabra et al. 2004a). Further studies are being carried out to determine if strains with different virulence inhibit NO the same way and the same way and the involvement of phosphatidylserine exposure in this evasion mechanism. The Brazilian contribution is and has been important to T. gondii biology. The many recent descriptions and genotyping of new isolates of this parasite in Brazil, specially in chickens (Dubey et al. 2008) shows that lot of work need to be done to better understand virulence to different worm blood animals and humans and how the parasite has disperse around the world. Thus, we are sure that in the next 100 years the Brazilian contribution to the biology of this important intracellular pathogen will still be relevant. REFERENCES
Copyright 2009 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc09027f1.jpg] [oc09027f5.jpg] [oc09027f3.jpg] [oc09027f8.jpg] [oc09027f2.jpg] [oc09027f9.jpg] [oc09027f10.jpg] [oc09027f7.jpg] [oc09027f6.jpg] [oc09027f4.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}