 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 104, No. 2, March, 2009, pp. 201-210 Advances in understanding immunity to Toxoplasma gondii Elia D Tait; Christopher A Hunter+ Department of Pathobiology,
School of Veterinary Medicine, University of Pennsylvania, Hill Pavilion, Rm
341, 380. South University Avenue, Philadelphia, PA, USA 19104 Received 10 October
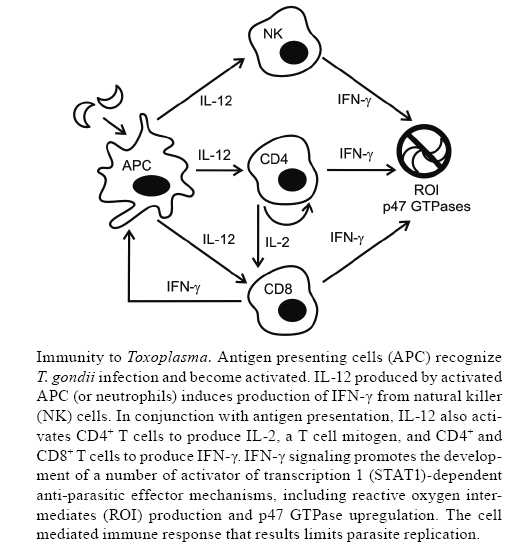
2008 Financial support: T32 AI007532, RO1 AI041158 Code Number: oc09036 ABSTRACT Toxoplasma gondii is an important cause of clinical disease in fetuses, infants and immunocompromised patients. Since the discovery of T. gondii 100 years ago, this pathogen and the host's immune response to toxoplasmosis have been studied intensely. This has led to the development of a working model of immunity to T. gondii, and has also resulted in fundamental new insights into the role of various cytokines in resistance to infection. By examining this organism, researchers have identified many of the requirements for resistance to intracellular pathogens and characterized numerous regulatory factors, including interleukin-10 (IL-10) and IL-27, which control inflammatory processes. In the next 100 years of T. gondii immunobiology, researchers will have the opportunity to answer some of the long-standing questions in the field using new techniques and reagents. These future studies will be vital in building a more comprehensive model of immunity to this pathogen and in advancing our understanding of immunoregulation, particularly in humans. Ultimately, the challenge will be to use this information to develop new vaccines and therapies to manage disease in affected patients. Key words:Toxoplasma gondii immunology - cytokines - microscopy - protozoan vaccines In 1907, at the Institut Pasteur in Tunis, while studying the infectious diseases of the desert rodent, Ctenodactylus gundii, Charles Nicolle observed the presence of a crescent-shaped organism in the tissues of these animals (Nicolle 1907). In 1908, Nicolle and his collaborator, Louis Manceaux, published the initial description of this discovery (Nicolle & Manceaux 1908). A year later, they provided a more complete report and named the organism Toxoplasma gondii, referencing the parasite's characteristic morphology and the source of the original isolate (Nicolle & Manceaux 1909). Concurrently, in Brazil, Alfonso Splendore (1908) identified the same microbe as a parasite of rabbits (Splendore 1908). At the time, the discovery of T. gondii perhaps seemed to be solely of academic interest; however, when Wolf et al. (1939) discovered the parasite in infants with encephalomyelitis, T. gondii was recognized as a cause of congenital disease. Since T. gondii's discovery, its clinical importance has influenced the research groups investigating the immunobiology of toxoplasmosis. Today, T. gondii is recognized as an important opportunistic pathogen of fetuses, newborns and patients with a variety of primary genetic and acquired immunodeficiencies (Petersen & Dubey 2001). The majority of immunocompromised patients that develop clinical disease have defects in T cell function, highlighting the importance of lymphocytes in controlling this persistent infection. Consequently, there has been a focus on understanding how T cells provide protection against disease and how cytokines modulate T cell responses (Lieberman & Hunter 2002). This review will provide an overview of these events, consider how the study of T. gondii has had a significant impact on the field of immunology and discuss future studies that may provide new insights into the mechanisms necessary for the control of this pathogen. The first 100 years: T. gondii immunobiology T. gondii is one of the world's most prevalent and successful parasites. Its biology is complex and numerous researchers worked throughout the 1960s and 1970s to elucidate the life cycle and means of transmission (Ajioka & Soldati 2007). This body of work is summarized elsewhere in this issue and here we focus on the immune responses that lead to the control of the asexual tachyzoite and bradyzoite forms. Intermediate hosts become infected with T. gondii after ingesting oocysts in the environment or tissue cysts from infected animals. Parasite replication in the intestine eventually leads to host cell lysis and parasite egress and tachyzoites disseminate throughout the host (Peter-sen & Dubey 2001). This process is poorly understood, but recent work suggests that CD11c+ dendritic cells (DC) may act as Trojan horses to spread the infection (Courret et al. 2006, Lambert et al. 2006). In immunodeficient hosts or during primary infection with highly virulent strains, the immune system cannot control parasite replication and clinical disease results (Fuentes et al. 2001). In most healthy hosts, the immune response controls replication, disease is limited, and physiological stress on the parasite causes tachyzoites, primarily in the brain and muscle tissue, to differentiate into bradyzoite cysts that persist throughout the host's life. Tissue cysts undergo periodic reactivation, but these events are controlled by an intact immune system. However, in chronically infected hosts that lose T cell function, reactivation may lead to disease (Petersen & Dubey 2001). The study of T. gondii immunobiology began in the 1940s, with Sabin and Feldman's (1948) description of complement-fixing antibodies to T. gondii in immune serum that led to the development of the Sabin-Feldman dye-test for sero-diagnosis. Subsequently, during the late 1960s, two vital components of the host immune response to T. gondii were uncovered. In 1968, Brownlee, Remington and others provided the first reports that T. gondii infection led to interferon production (Freshman et al. 1966, Remington & Merigan 1968, Rytel & Jones 1968). In 1967, Frenkel described the role of cell-mediated immunity in protection against this pathogen in the Golden hamster (Frenkel 1967) and, in 1972, Remington et al. established the importance of macrophages in controlling replication of T. gondii. These studies laid the groundwork for the next 40 years of work in the field, in which researchers focused on the cytokine and cell-mediated components of immunity required for resistance to this organism. During the 1970s and 1980s, the population of immunocompromised hosts grew dramatically and the incidence of opportunistic infections increased. Transplant and cancer patients on newly-developed immunosuppressive drugs, and patients with immunosuppressive cancers, had an increased incidence of toxoplasmosis caused by parasite reactivation in the latently infected host (Remington 1974). In the early 1980s, with the emergence of the human immunodeficiency virus (HIV) and acquired immune deficiency syndrome (AIDS), a new cohort of immunosuppressed patients deficient in CD4+ T cells was identified. These patients were susceptible to a variety of intracellular pathogens, including T. gondii, and presented with symptoms of toxoplasmic encephalitis (TE) following parasite reactivation (Horowitz et al. 1983, Pitchenik et al. 1983, Luft et al. 1984, Levy et al. 1988, Jones et al. 1996). These clinical observations were corroborated experimentally in studies in the mouse model, in which depletion of CD4+ or CD8+ T cells during chronic infection resulted in TE (Vollmer et al. 1987, Suzuki & Remington 1988, Gazzinelli et al. 1991, 1992b, Parker et al. 1991). Together, these clinical observations and laboratory studies provided evidence for the vital role for CD4+ and CD8+ T cells in the long-term control of T. gondii. Following the initial reports in the late 1960s that described interferon production in response to T. gondii, cellular immunity and cytokine production were linked in 1983 when Rubin et al. reported that interferon-γ (IFN-γ) produced by T lymphocytes activated macrophages to produce reactive oxygen intermediates (ROI) and kill T. gondii (Nathan et al. 1983). By the late 1980s, IFN-γ produced by CD4+ and CD8+ T cells was identified as the major mediator of protection against T. gondii (Suzuki et al. 1988, 1989) and there were suggestions that accessory cell products were responsible for inducing IFN-γ production from T lymphocytes and natural killer (NK) cells (Kelly et al. 1989). At the same time, Trinchieri and colleagues had identified the heterodimeric cytokine interleukin-12 (IL-12) as a "powerful immunopotentiating agent" (Kobayashi et al. 1989, Chan et al. 1991) that stimulated IFN-γ production from human and murine T lymphocytes and NK cells. The Sher, Kasper and Remington laboratories established that IFN-γ production in response to T. gondii was largely IL-12-dependent in both immunodeficient and immunocompetent mice, with important roles for IL-2 and tumor necrosis factor-α (TNF-α) as co-factors (Gazzinelli et al. 1993, 1994a, b, Hunter et al. 1994, 1995, Khan et al. 1994). Macrophages (Gazzinelli et al. 1993), DC (Reis e Sousa et al. 1997, Liu et al. 2006) and neutrophils (Bliss et al. 1999) were all identified as sources of IL-12 during toxoplasmosis, although DC, including conventional CD8+ (Reis e Sousa et al. 1997) and plasmacytoid DC (Pepper et al. 2008), now appear to be the major contributors (Liu et al. 2006). These different cell types become activated and produce IL-12 following the engagement of either chemokine (C-C motif) receptor 5 (CCR5) (Aliberti et al. 2000) or Toll-like receptors (TLR), including TLR 2 (Mun et al. 2003) and TLR 11 (Yarovinsky et al. 2005) by parasite-derived molecules. In recent years, the study of IFN-γ-dependent effector mechanisms that limit parasite replication have underlined the importance of the IL-12/IFN-γ axis. IFN-γ signals through signal transducer and activator of transcription 1 (STAT1) to activate a variety of antimicrobial effector mechanisms, including the upregulation of inducible nitric oxide synthase (iNOS). ROI produced by iNOS were initially thought to be the host's primary means of controlling parasite replication, as illustrated by the increased susceptibility of iNOS knockout mice. However, unlike IFN-γ knockout mice, iNOS knockout mice survive the acute phase of infection and do not succumb until the chronic phase. These data indicated the presence of IFN-γ-dependent, iNOS-independent pathways to inhibit parasite growth (Scharton-Kersten et al. 1997). One such pathway includes the activation of the family of p47 guanosine triphosphatases (GTPases), which are upregulated in response to IFN-γ (Taylor et al. 2000, Collazo et al. 2001). Studies utilizing specific p47 GTPase knockout animals have provided evidence that these proteins degrade the parasitophorous vacuole in T. gondii-infected cells and are involved in the process of autophagy (Martens et al. 2005). Ultimately, though mice deficient in STAT1 (Gavrilescu et al. 2004, Lieberman et al. 2004), iNOS (Scharton-Kersten et al. 1997) and members of the 47 kD GTPase family (Taylor et al. 2000, Collazo et al. 2001) all show increased susceptibility to T. gondii infection, we still do not fully understand the mechanisms by which IFN-γ mediates protection. Ongoing studies seek to better characterize IFN-γ-dependent anti-parasitic effector mechanisms. Taken together, the studies reviewed here, and others, have led to the development of a model of immunity to T. gondii, summarized in the Figure. Accessory cells, notably DC, recognize that the host is infected with T. gondii, likely through pattern recognition receptors. These cells become activated and produce IL-12, which promotes NK cell production of IFN-γ. IL-12 derived from antigen presenting cells (APC), in the context of antigen presentation, also drives CD4+ and CD8+ T cell activation and IFN-γ production. In addition, activated CD4+ T cells produce IL-2, an important T cell mitogen. Together, these events result in the convergence of large numbers of parasite-specific CD4+ and CD8+ T cells that produce IFN-γ at sites of parasite invasion. IFN-γ signaling, in a STAT1-dependent pathway, leads to the generation of anti-parasitic effector mechanisms, including the production of ROI and the activation of the p47 GTPases. Immunoregulation during T. gondii infection The study of the immune response to parasites has consistently provided new insights into the importance of cell-mediated immunity and the role of cytokines in the control of intracellular pathogens. For example, the study of the interactions between T. gondii and the host has provided a better understanding of how an immune response is initiated, expanded, and contracted to limit pathogen growth and replication while controlling immunopathology. Many mechanisms have been identified that modulate infection-induced pathology, including the activity of lipoxins, IL-10 and IL-27 (Lieberman & Hunter 2002, Aliberti 2005). Two of the most important anti-inflammatory cytokines, IL-10 and IL-27, have been studied extensively during toxoplasmosis and this work has contributed to the basic understanding of the function of these factors. IL-10 limits infection-induced immune pathology - In 1989, a new cytokine, initially called cytokine synthesis inhibitory factor, and later designated as IL-10, was identified based on its ability to antagonize T helper 1 (Th1) responses (Fiorentino et al. 1989). Today, IL-10 is considered to be an inhibitor of Th1, Th2 and Th17 immune responses (Moore et al. 2001, Lieberman & Hunter 2002, O'Garra & Vieira 2007). It is produced by macrophages, monocytes, DC, B cells, and CD4+ and CD8+ T cells, and acts broadly on accessory cells to downregulate proinflammatory cytokine production and major histocompatibility complex (MHC) and costimulatory molecule expression. Thus, T cell priming and activation are dampened primarily through IL-10's effects on accessory cells, although there are reports that IL-10 may also directly suppress CD4+ T cell proliferation and cytokine production (Couper et al. 2008). In 1992, the Sher laboratory reported, consistent with IL-10's inhibitory effects on macrophages, that IL-10 antagonized the ability of IFN-γ-primed macrophages to kill intracellular T. gondii (Gazzinelli et al. 1992a). Also at this time, IL-10 was identified as a factor induced by T. gondii infection (Gazzinelli et al. 1992a, 1994a, Hunter et al. 1993, Burke et al. 1994) that contributed to the suppression of T cell function (Candolfi et al. 1995, Khan et al. 1995). These findings contributed to a model in which T. gondii induces IL-10 production to limit the host immune response and favor parasite replication. Unexpectedly, IL-10-/- mice infected with T. gondii succumbed to systemic inflammatory disease (Gazzinelli et al. 1996, Neyer et al. 1997) mediated by CD4+ T cells (Gazzinelli et al. 1996, Suzuki et al. 2000, Wilson et al. 2005) and dependent on CD40-CD40L and CD28-CD80/CD86 interactions (Villegas et al. 2000, Wille et al. 2002). Thus, IL-10 was established as a vital player in the control of immunopathology during toxoplasmosis. Given the importance of IL-10 in preventing and limiting inflammation, identifying the source of this cytokine has become an important issue. In 2007, Sher et al. identified CD4+Tbet+Foxp3- Th1 cells that produced large amounts of IFN-γ and were also potent sources of protective IL-10 (Jankovic et al. 2007). These cells do not appear to be regulatory T cells, but rather effectors that also possess the capacity to produce IL-10 and limit immune pathology. This study, and others that found similar results in different systems, indicates that autoregulatory mechanisms that promote effector CD4+ T cell production of IL-10 have a key role in maintaining balanced, yet efficacious, immune responses to T. gondii and other pathogens (O'Garra & Vieira 2007). Though the importance of IL-10 in T. gondii infection is clear, questions remain regarding how IL-10 integrates with other immunoregulatory pathways. Though CD4+ T cells are an important source of IL-10 during toxoplasmosis, there are other cellular sources of IL-10 during acute and chronic infection and their relative contribution to the control of immune-mediated pathology is unclear. Additionally, how these cytokine-producing cells migrate and behave in secondary lymphoid organs versus sites of tissue inflammation is unknown. In order to address these questions, new approaches and reagents are being developed, including intravital imaging and IL-10 reporter mice and these advances will provide opportunities to better understand this regulatory cytokine. IL-27: a new anti-inflammatory cytokine - In 2002, researchers at DNAX described a heterodimeric cytokine, IL-27, composed of the p28 and Epstein Barr virus-induced gene 3 (EBI3) subunits. The p28 subunit was identified in a computational screen designed to identify homologs of the IL-6/IL-12 family of cytokines, and EBI3 was identified as a p28 binding partner in immunoprecipitation and secretion studies (Pflanz et al. 2002). Initial findings indicated that IL-27 was produced by APC, and acted on naïve T cells to induce proliferation and Th1 polarization (Pflanz et al. 2002). These effects were mediated by IL-27 binding its receptor, composed of IL-27 receptor α (WSX-1) and glycoprotein 130, and signaling through STAT1 to induce expression of T-bet and IL-12 receptor β2 (Hibbert et al. 2003, Lucas et al. 2003, Takeda et al. 2003, Villarino et al. 2003, Pflanz et al. 2004). Although the initial characterization suggested that IL-27 had proinflammatory properties, when WSX-1 deficient mice were challenged with T. gondii, they succumbed to infection and had unexpectedly low parasite burdens and significant immunopathology. These mice had high levels of Th1 cytokines, increased T cell activation and proliferation, and cellular infiltrates and tissue destruction in the liver and lungs. The immune pathology noted was dependent upon the presence of CD4+ T cells, demonstrating a phenotype similar to that observed in IL-10 deficient animals; however, overall IL-10 levels were not decreased in acutely infected WSX-1 deficient animals. In vitro studies indicated that IL-27 was only proinflammatory in conditions in which IL-12 was limiting, whereas in conditions where IL-12 was abundant (as in toxoplasmosis), IL-27 actually played a suppressive role (Villarino et al. 2003). Subsequent studies in other models of infectious and autoimmune disease have confirmed an anti-inflammatory role for IL-27 in Th1 (Rosas et al. 2006, Wirtz et al. 2006, Sonoda et al. 2007), Th2 (Artis et al. 2004, Miyazaki et al. 2005, Shimizu et al. 2005) and Th17 responses (Batten et al. 2006, Stumhofer et al. 2006, Fitzgerald et al. 2007), and it can directly induce IL-10 production (Awasthi et al. 2007, Fitzgerald et al. 2007, Stumhofer et al. 2007). The study of IL-27's role during toxoplasmosis has yielded important insights into its biological activities, as well as emphasizing the importance of immunoregulation during infection. However, the function and regulation of IL-27 during T. gondii infection is still poorly understood. The cellular sources of IL-27, it's role during chronic versus acute infection, the contributions of the p28 and EBI3 subunits, and how it intersects with other cytokine signaling networks, including the importance of the EBI3 subunit in IL-35 signaling (Collison et al. 2007), are open questions. As has been the case for IL-10, the study of IL-27 during toxoplasmosis promises to further our understanding of T. gondii immunobiology and the function of IL-27. The next 100 years: new challenges in T. gondii immunobiology In the 100 years since the discovery of T. gondii, we have developed a complex picture of the events vital for controlling toxoplasmosis. This area of study has consistently been at the forefront of immunology and researchers are taking advantage of new culture techniques, immunological assays and animal models, including knockout and transgenic mice, to better understand how the host is able to recognize and control this pathogen. While there has been enormous progress in this field, many questions remain. Particularly, the capacity of cytokines to regulate T cell responses in vivo is not fully understood and the events that occur during the human immune response to toxoplasmosis are unclear. The following section will provide a personal perspective on some of the questions and areas of investigation that will help address these specific questions. Imaging the immune response to T. gondii - Historically, in order to examine immune responses at the cellular level, immunologists have relied upon techniques and reagents that provided "snapshots" of lymphocyte behavior at a given time point, long after early activation or priming events had occurred (Mempel et al. 2004, Sumen et al. 2004, Germain et al. 2005, 2006, Garside & Brewer 2008). Some of the outstanding questions in the field of T. gondii immunology involve understanding how cytokine-producing accessory cells and T cells interact to coordinate a protective immune response and, subsequently, how effector T cells interact with T. gondii-infected cells to control parasite replication. Addressing these issues will require a more dynamic approach to analyzing cellular immunity, in which cell-cell interactions in vivo can be observed in real-time. In the past decade, advances in whole-body animal imaging and intravital multiphoton (MP) microscopy have provided new tools to observe and study the behavior of single cells. Here we will provide examples of questions in the field that have been addressed using imaging technology and discuss how these studies can be extended to further investigate the behavior of individual immune cells during toxoplasmosis. Until recently, we have had a limited insight into how T. gondii-infected cells behave in vivo during toxoplasmosis and how T cells and APC interact with infected cells. The development of transgenic parasite lines that express fluorescent (Striepen et al. 1998) or bioluminescent proteins has allowed for imaging and flow cytometry studies that analyze infected cells in vivo and in vitro. Using parasites that express luciferase in conjunction with whole-body animal imaging techniques, patterns of T. gondii dissemination (Hitziger et al. 2005, Saeij et al. 2005), reactivation (Dellacasa-Lindberg et al. 2007) and stage conversion (Saeij et al. 2008) in vivo have been described. Fluorescent parasites have been used to characterize CD11c+ and CD11b+ cells as Trojan horses that spread T. gondii throughout the host (Courret et al. 2006, Lambert et al. 2006), investigate apoptotic and cytolytic pathways in infected cells (Persson et al. 2007), describe how infected cells present T. gondii antigen (Gubbels et al. 2005, Dzierzsinski et al. 2007) and examine interactions between parasites and DC (Boes et al. 2002, McKee et al. 2004). Additionally, transgenic parasite lines that express a model antigen, such as ovalbumin (Pepper et al. 2004, Gubbels et al. 2005), β-galactosidase (Kwok et al. 2003) or Eα (Pepper et al. 2008), allow antigen-specific T cell responses to be tracked. Using bioluminescent and fluorescent parasites in conjunction with model antigen systems, the interaction between infected target cells and antigen-specific T cells can be investigated using existing technologies (Dzierszinski & Hunter 2008) and, ultimately, as resolution improves, it may be possible to visualize these interactions using whole-body imaging. While multiple groups have identified important roles for inflammatory and immunoregulatory cyto-kines during toxoplasmosis, additional studies will be required to identify the cellular sources of these factors and determine when and where they are produced. To date, the tools to investigate these issues have had limitations. Ex vivo cytokine production has been assayed in response to a stimulus, using intracellular cytokine staining or enzyme-linked immunoassay as read-outs, but stimulating cells in this manner may not reflect the events that occur in vivo (Mohrs et al. 2001). An alternative approach to studying these cells is through the use of cytokine-reporter mice, in which an appropriate cytokine promoter drives the expression of a fluorescent protein. Today, there are reporters available for multiple cytokines, including IFN-γ (Stetson et al. 2003), IL-4 (Mohrs et al. 2001), IL-10 and IL-12p40 (Reinhardt et al. 2006). Thus far, these reporters have been used primarily in flow-based assays to detect cytokine-producing cells. However, these animals lend themselves well to imaging studies, including MP microscopy, and could be applied to investigate real-time immune responses to T. gondii. In the past decade, advances in the field of MP microscopy have allowed researchers to image deep into tissues for long periods of time to accurately monitor and quantitate lymphocyte behavior and migration in vivo. MP microscopy uses pulses of low-energy, high-density photons to excite fluorophors, allowing for four-dimensional analysis of immune cells with minimal photo-bleaching. Intravital MP microscopy, in conjunction with the development of new fluorescent reagents and surgical techniques, has led to the development of novel questions and hypotheses (reviewed in Mempel et al. 2004, Sumen et al. 2004, Germain et al. 2005, 2006, Garside & Brewer 2008). Cytokine reporter mice will provide new insights into how cytokine-producing cells migrate and interact with other cells. This work will be important in understanding the broad orchestration of cytokine production throughout an immune response and the detailed role of the cytokine-producer in distinct microenvironments. In addition to examining the migration and behavior of cytokine-producing cells, other issues in T. gondii immunology can be addressed using MP microscopy. These include determining what interactions T cells and APC participate in, and how T cells, APC and other cell types migrate and behave during primary infection (Chtanova et al. 2008). Imaging studies using reporter mice for lineage-specific cell surface markers can be used to quantitate the location, frequency and duration of encounters between T cells and various APC. This information will give us a better understanding of how effector T cells are primed and activated and whether migration within lymphoid tissues and through sites of infection differs. Notably, the nature of lymphocyte behavior, migration and cell-cell interactions in the brains of chronically infected mice is largely unknown. Intravital MP imaging will be key in developing a model of the cellular immune response in this tissue site. Human immunology - While toxoplasmosis has been recognized as an important human disease for almost 75 years, the rodent has been the model of choice to study the immune response to T. gondii. Rodents are natural hosts of this organism, are easy to use and maintain, can be genetically manipulated to produce knockouts and transgenics and there is a wide array of reagents to study the murine immune system. Additionally, the technical and ethical difficulties of human research have presented a substantial challenge to researchers with an interest in better understanding how the human host interacts with the parasite. Thus, with notable exceptions, we know little about the human immune response to toxoplasmosis. While most immunocompetent humans that become infected with T. gondii are asymptomatic, the parasite can cause disease in two main groups of people: those with acquired or primary immunodeficiencies and fetuses infected in utero (Petersen & Dubey 2001). These two classes of patients present different challenges to health professionals in terms of diagnosis, case management and drug treatment. For basic researchers, studying how and why disease occurs in susceptible patients may provide the opportunity to develop new vaccines and immune-based therapies. Congenital toxoplasmosis occurs in infants that are infected during gestation, following a primary challenge of the mother (Wong & Remington 1994). Interestingly, not all pregnant women that contract a primary infection transmit the parasite to their fetus. The mechanisms that influence vertical transmission are poorly defined, although there is a positive correlation between rate of transmission and infection during the second or third trimester of pregnancy (Desmonts & Couvreur 1984, Dunn et al. 1999). The placenta itself also appears to play an important role in mediating transmission (Pfaff et al. 2007) and studies in the murine model suggest a role for IFN-γ in facilitating transmission (Abou-Bacar et al. 2004). Additionally, there is evidence that fetuses infected in utero may be tolerized to T. gondii antigens in both the mouse (Suzuki & Kobayashi 1990) and the human (McLeod et al. 1985, Hara et al. 1996, Yamamoto et al. 2000). However, the nature of the fetal response to T. gondiii challenge, and how this response ultimately impacts the long-term consequences of in utero infection, is unclear. One of the most important challenges in managing congenital toxoplasmosis is developing effective preventative measures. In mothers previously exposed to T. gondii, the fetus is very rarely infected (Remington et al. 2000), suggesting that natural maternal immunity to T. gondii is sufficient to protect the fetus from vertical transmission. In principle, this protection could be mimicked using a vaccine. Numerous strategies have been employed to elicit protective immunity in mouse models of congenital toxoplasmosis, including recombinant protein (Letscher-Bru et al. 2003) and DNA-based vaccine strategies (Mévélec et al. 2005), but to date, a human vaccine has not been developed. In the immunocompetent murine host during secondary challenge, CD8+ T cells are vital in mediating protection (Gazzinelli et al. 1991). This observation suggests that a successful vaccine should mobilize parasite-specific CD8+ T cells to prevent primary infection (Pfaff et al. 2007). Unfortunately, the study of antigen-specific T cell responses in humans has been limited due to reagent availability and thus the evaluation of human cellular responses to vaccines has been difficult. In the past decade however, numerous groups have developed MHC class I tetramer reagents to identify and study antigen-specific CD8+ (Serbina & Pamer 2003) and CD4+ (Mallone & Nepom 2004) T cells in human viral and bacterial infections. These reagents specifically identify T cells with the T cell receptor (TCR) for a peptide of interest and have been used to track vaccine-specific responses in humans (Serbina & Pamer 2003). Recently, a number of groups have collaborated to develop tetramer reagents for immundominant T. gondii CD8+ T cell epitopes in the murine model (Blanchard et al. 2008, Frickel et al. 2008). This approach could be applied to develop tetramers for use in human studies, so that the T. gondii-specific T cell responses of newborns infected in utero and vaccinated women before and during pregnancy could be examined. The development of these tetramer reagents would allow for the detailed investigation of fetal anergy in response to T. gondii antigens and would provide a read-out of vaccine efficacy. Patients with primary or acquired immunodeficiencies represent a broad group of individuals with various deficits in T cell, monocyte, cytokine and B cell function. Cancer patients with immunosuppressive cancers, including leukemia and lymphoma, experience recrudescence of chronic infection as T cell function declines. Cancer or transplant patients receiving immunosuppressive drugs as part of a chemotherapeutic regimen also present with reactivated toxoplasmosis (Armstrong et al. 1971, Ruskin & Remington 1976, Frenkel et al. 1978, Hakes & Armstrong 1983, Derouin et al. 1992, Israelski & Remington 1993, Lappaleinen et al. 1998). Patients with hyper-IgM syndrome, in which the CD40/CD40L interaction is disrupted, resulting in defects in cell-mediated immunity and an inability to class switch (Aversa et al. 1994), are susceptible to primary infection (Subauste et al. 1999). Finally, HIV-infected patients, as described above, have an increased incidence of TE (Horowitz et al. 1983, Pitchenik et al. 1983, Luft et al. 1984, Levy et al. 1988, Jones et al. 1996). Widespread use of highly active anti-retroviral therapy (HAART) has led to better control of viral replication, accompanied by an increase in immune function; thus, TE and other opportunistic infections are not as prevalent in these patients as was once the case. However, new immunosuppressed cohorts are continually emerging, including patients with autoimmune disorders being treated with immunmodulatory drugs (Hemmer et al. 2006), and growing drug resistance to HAART may result in the reemergence of TE as a clinical problem in HIV/AIDS patients (Alfonso et al. 2002, Wensing & Boucher 2003). Together, these at-risk patient groups present challenges for basic human immunology research and clinical management. Patients with increased susceptibility to toxoplasmosis provide the researcher with the opportunity to better understand how the human immune system functions. Importantly, these patient cohorts highlight the need for innovative new drug treatments and effective therapies (Fischer 2007). With new advances in human immunology, including the application of multi-color flow cytometry for detection of cell surface markers, cytokine expression and intracellular cell signaling molecules, researchers and clinicians will be able to develop a better understanding of human immunity to T. gondiii and will also have a greater ability to assess the efficacy of treatment. These tools also promise to be useful in identifying and developing immunomodulators and immune therapies to manage this infection. Since the intact human immune response controls parasite replication effectively in most cases, immune therapy to replace or augment dysfunctional immune responses in patients with immunodeficiencies may be an effective approach to managing this condition (McCabe 2001). In the first 100 years of research in T. gondii immunology, we have developed an initial model to understand the complexity of the immunoregulatory networks that limit immunopathology and control infection. The study of T. gondii has made significant contributions to the field of immunology and is well-suited to continue to do so, in understanding cytokine function and investigating human immune responses. Hopefully, in the next 100 years of T. gondii research, investigators will be able to better understand how the innate and adaptive immune systems are coordinated and controlled in mice and humans, develop a protective vaccine to prevent congenital toxoplasmosis and generate new effective treatment regimens for patients affected by toxoplasmosis. The greatest challenge will be to intelligently question our current models for understanding the immune response to T. gondii and creatively test these models to move the field forward. REFERENCES
Copyright 2009 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc09036f1.jpg] |
| |||||||||

{kind=link}