 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 104, No. 3, May, 2009, pp. 411-418 ARTICLES Evaluation of an enzyme-linked immunoelectrotransfer blot test for the confirmatory serodiagnosis of human toxocariasis William H Roldán+; Yrma A Espinoza Sección
de Parasitología, Instituto de Medicina Tropical Daniel A. Carrión,
Facultad de Medicina, Universidad Nacional Mayor de San Marcos, Calle José
Santos Chocano 199, CP 10138, Callao, Perú Received 4 May
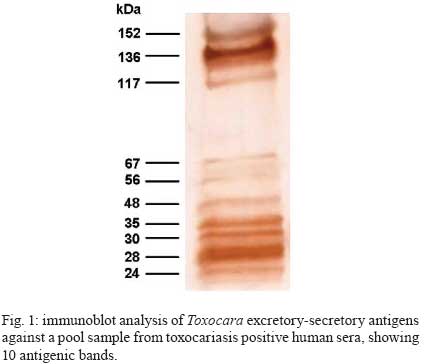
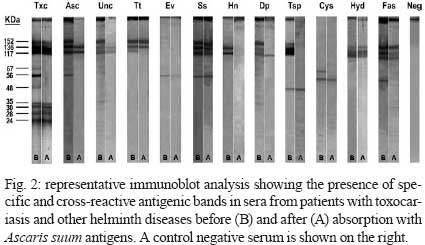
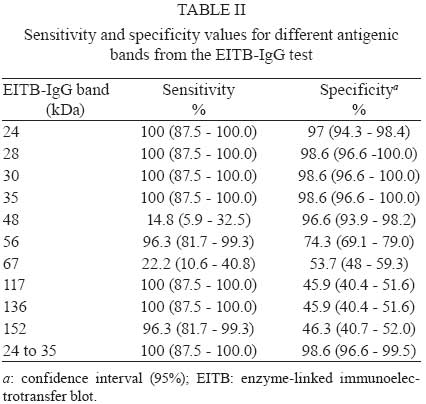
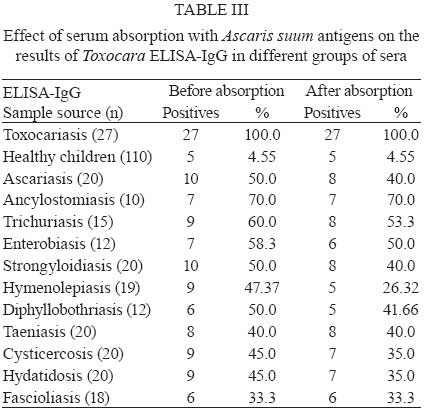
2008 Code Number: oc09065 ABSTRACT To improve the serodiagnosis of human toxocariasis, a sensitive and specific enzyme-linked immunoelectrotransfer blot (EITB-IgG) test was developed and evaluated using Toxocara canislarvae excretory-secretory antigens for detecting anti-Toxocara IgG antibodies. The EITB-IgG profile of toxocariasis was characterized by comparing 27 sera from patients with toxocariasis, 110 sera from healthy subjects and 186 sera from patients with other helminth diseases (ascariasis, ancylostomiasis, trichuriasis, enterobiasis, strongyloidiasis, hymenolepiasis, diphyllobothriasis, taeniasis, cysticercosis, hydatidosis and fascioliasis). Antigenic bands of 24, 28, 30, 35, 56, 117, 136 and 152 kDa were predominantly recognized in sera from all patients with toxocariasis. However, only bands of 24-35 kDa were highly specific for Toxocara infection (98.3%), whereas other antigenic bands observed displayed cross-reactivity. Additionally, when the results of the EITB-IgG test were compared to those of the ELISA-IgG test, a 100% concordance was observed for positive results in human toxocariasis cases. The concordance for negative results between the two tests for healthy subjects and patients with other helminth diseases were 96.3% and 53.7%, respectively, showing that the EITB-IgG test has a higher specificity than ELISA. In conclusion, the EITB-IgG test is a very useful tool to confirm the serological diagnosis of human toxocariasis. Key words:Toxocara - toxocariasis - immunodiagnosis - cross-reactions - EITB Human toxocariasis is a major parasitic zoonosis, caused by infection with the larvae of Toxocara canis, the common roundworm of dogs, and less frequently, of Toxocara cati, the roundworm of cats (Schantz & Glickman 1983, Despommier 2003). The incidence of human toxocariasis is unknown because it is a non-communicable disease in many countries. However, many cases of this disease have been reported around the world (Glickman & Schantz 1981, Despommier 2003). Humans become infected by ingestion of embryonated Toxocara eggs found in soil, contaminated vegetables, on dirty hands or in undercooked giblets. These eggs hatch in the proximal intestine and the released larvae penetrate the mucosa and migrate through the inner organs, causing an inflammatory response (Magnaval et al. 2001). Toxocara cannot complete its life cycle in humans and parasite development is arrested at the larval stage. The migrating larvae give rise to the clinical syndromes of visceral larva migrans (VLM), ocular toxocariasis (OT) and a non-symptomatic infection [covert toxocariasis (CT)] (Magnaval et al. 2001). VLM includes fever, abdominal pain, malaise, weight loss, skin rash, hepatomegaly, hypergammaglobulinaemia and respiratory symptoms associated with eosinophilia (Magnaval et al. 2001). Childhood bronchial asthma has also been associated with toxocariasis (Oteifa et al. 1998, Chan et al. 2001). OT, caused by larvae migration to the eye, can present as strabismus, pars planitis, endophthalmitis, uveitis, retinal granuloma and retinal detachment, leading to vision loss (Gillespie et al. 1993). CT presents with a wide range of nonspecific symptoms, including cough, abdominal pain, headache and sleep and behavioral disturbances (Taylor et al. 1988). Since the parasitological diagnosis of this infection is hampered by the impossibility of detecting larvae in tissues, the diagnosis of toxocariasis is based on clinical and serological data. The test commonly employed for the serodiagnosis of toxocariasis is ELISA using Toxocara excretory-secretory (TES) antigens to detect serum anti-Toxocara IgG antibodies (De Savigny et al. 1979, Jacquier et al. 1991). However, some components of TES antigens are not specific, demonstrating moderate cross-reactivity with other helminth species (Speiser & Gottstein 1984, Lynch et al. 1988, Jacquier et al. 1991, Nunes et al. 1997). Sera from patients with ascariasis, filariasis, schistosomiasis and strongyloidiasis also react with TES antigens (Maizels et al. 1984, Nunes et al. 1999, Yamasaki et al. 2000). In northern countries, the TES antigens are more specific for serodiagnosis of toxocariasis, but the level of specificity is unsatisfactory in countries where co-infections with other helminths are frequent. Thus, in the tropics, Toxocara serodiagnosis remains a problem. In order to reduce the possibility of cross-reactions with other parasites, many authors have proposed that serum absorption with extracts of a wide variety of non-homologous parasites be used (Lynch et al. 1988, Camargo et al. 1992, Nunes et al. 1997), but as the availability of these parasites is limited, their preparation is often difficult. Enzyme-linked immunoelectrotransfer blot (EITB) or immunoblot is a test that combines the high sensitivity of the immunoenzymatic tests with the high resolution of sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE); this method has been successfully adapted for the confirmatory serodiagnosis of various parasitic diseases in humans, such as schistosomiasis, hydatidosis, cysticercosis, taeniasis, fascioliasis and strongyloidiasis (Tsang et al. 1989, Hillyer et al. 1992, Verastegui et al. 1992, Valli et al. 1999, Wilkins et al. 1999, Silva et al. 2003). Western blotting for serodiagnosis of human toxocariasis has also been described (Magnaval et al. 1991). Human helminthiasis is very frequent in various regions of Peru (Gilman et al. 1991, Marcos et al. 2005, Larocque et al. 2006, Romani et al. 2006, Yori et al. 2006, Chero et al. 2007, Cleary et al. 2007, García et al. 2007) and cross-reactions with these parasitic infections would be enough to cause confusion in the serodiagnosis of toxocariasis. Although the lack of specificity of the ELISA test for the diagnosis of human toxocariasis can be reduced by using serum pre-absorbed with regional helminth antigens (Lynch et al. 1988, Camargo et al. 1992, Nunes et al. 1997), we herein propose that it would be useful to confirm any positive results from the ELISA test (where pre-absorption is not carried out) using the EITB test on patients with suspected toxocariasis. In this study, we describe the development of an EITB test for the confirmatory serodiagnosis of human toxocariasis and compare the outputs of this assay with those of the ELISA test. PATIENTS, MATERIAL AND METHODS Patients and sera - Twenty seven serum samples from patients with toxocariasis diagnosed based on clinical (ocular or visceral forms of the disease), hematological and serological findings, were used to identify specific Toxocara IgG antibodies. Serum samples from 110 healthy children (with repetitive negative results for intestinal parasite examination) were used as controls. In order to assess cross-reactivity, serum samples from 186 patients with parasitological or serological confirmed helminthiasis, including ascariasis (20 cases), ancylostomiasis (10 cases), trichuriasis (15 cases), enterobiasis (12 cases), strongyloidiasis (20 cases), hymenolepiasis (19 cases), diphyllobothriasis (12 cases), taeniasis (20 cases), cysticercosis (20 cases), hydatidosis (20 cases) and fascioliasis (18 cases), were used. Obtaining fully embryonated T. canis eggs - T. canis eggs were obtained by dissection of gravid adult female worms and incubated for 30 days at 28°C in a solution containing 2% formalin and 1% sodium hypochlorite for embryonation. The embryonated eggs were washed repeatedly with sterile distilled water and de-coated for 20 min in a 5% sodium hypochlorite solution at 37°C (Espinoza et al. 2003). The egg suspension was then washed repeatedly in sterile distilled water until all traces of chlorine had been removed. TES antigens - Fully embryonated and de-coated T. canis eggs were hatched by mechanical treatment with sterile glass beads for 15 min and the larvae were collected in a Baermann's apparatus (Lescano 1991). The TES antigens were produced according to De Savigny (1975), modified by Bowman et al. (1987). The supernatant from Toxocara larvae cultures, in RPMI 1640 medium supplemented with HEPES and glutamine (Sigma-Aldrich Co, Saint Louis, USA), was removed weekly, then dialyzed against distilled water and concentrated using PEG 20,000 (mol. wt) (Sigma-Aldrich Co, Saint Louis, USA). In order to inactivate any enzymatic activity of the TES antigens, Phenylmethylsulfonyl fluoride (Sigma-Aldrich Co, Saint Louis, USA) was added to a final concentration of 0.005 M. The protein content of the TES antigens was estimated using the Lowry method (Lowry et al. 1951) and the samples were kept in aliquots at - 20°C until use. Ascaris suum worm extract - This antigenic extract was obtained via a previously described method (Camargo et al. 1992). Protein concentrations of this antigenic extract samples were estimated (Lowry et al. 1951) and the samples were kept in aliquots at 20°C until use. Serum absorption with A. suum worm extract - In order to assess the effect of the previous serum absorption with this heterologous antigenic extract on the results in both tests, each human serum to be assayed was pre-absorbed (v/v) with soluble A. suum extract at a final concentration of 60 µg/ml of antigen. After incubation at 37°C for 1 h, dilutions of the absorbed sera were made and added to the nitrocellulose (NC) strips or microtitration wells. All the serum samples were re-tested twice to confirm the effect of the absorption procedure with A. suum antigens. EITB-IgG - The TES antigens were electrophoretically separated and transferred to NC membranes (Sigma-Aldrich Co, Saint Louis, USA, NC) according to the procedures described by Tsang et al. (1991) with some modifications. The TES antigens were diluted to a final concentration of 200 μg/mL with sample buffer (2.5 mM Tris-HCl, pH 8.0, containing 1% SDS, 50 mM dithiothreitol, 0.4% glycerol and 0.025% bromophenol blue), and then heated at 65°C for 15 min. The TES proteins were separated by SDS-PAGE in a 4-16% resolving gradient gel using a Miniprotean III system (Bio-Rad, Hercules, USA). Samples of TES were electrophoresed with constant current of 5 mA for 15 min and then 25 mA, until the bromophenol blue reached the end of the gel. The relative molecular weight (MW) was calculated using the wide range Sigma-MarkerTM MW standard (6.5 - 200 kDa, Sigma-Aldrich Co, Saint Louis, USA). The separated TES proteins were transferred to a 0.2 μm pore size, 8 x 8 cm NC sheets using a Trans-blot Cell system (Bio-Rad, Hercules, USA) with a constant current of 2.0 A for 2 h. The NC sheets were washed for 30 min with 0.01 M phosphate-buffered saline-0.3% Tween 20 (PBS-T), cut into 3-mm-wide strips and stored at - 20°C until use. NC strips were incubated with human serum samples diluted at 1:100 in PBS-T containing 5% non-fat milk (Nestlé Carnation, USA) overnight at 4°C. After washing three times for 5 min with PBS-T, the strips were incubated for 3 h at RT with an anti-human IgG peroxidase conjugate (Sigma-Aldrich Co, Saint Louis, USA) diluted 1:2000 in PBS-T. After washing with PBS-T, the strips were developed using a freshly prepared substrate solution containing 15 mg of 3,3' -diaminobenzidine tetrahydrochloride (Sigma-Aldrich Co, Saint Louis, USA), 30 mL of PBS and 10 μL of 30% hydrogen peroxide. The reaction was stopped after 5 min by washing with tap water. Positive reactions were determined by the visualization of defined brown bands judged using the naked eye. ELISA-IgG - Standard ELISA was performed according to the procedures described by Roldán et al. (2006) with some modifications. Briefly, 96-well flat-bottom polystyrene microtitration plates (Thermo Scientific Inc, USA) were coated with TES antigens (0.63 μg/mL) in 0.05 M carbonate-bicarbonate buffer, pH 9.6 (100 ¼L per well) overnight at 4ºC. All incubation volumes were 100 μL per well. The microtiter plates were washed with PBS-0.1% Tween 20 (PBS/T) and then nonspecific reactions were blocked for 2 h with PBS/T containing 5% non-fat milk (Nestlé Carnation, USA). All sera (diluted 1:200 in blocking solution) were added and incubated for 60 min at RT. After three washes with PBS/T, peroxidase conjugate anti-human IgG (Sigma-Aldrich Co, Saint Louis, USA) was added at a dilution of 1:4000 and the plates incubated for 60 min at RT. After three washes with PBS/T, the plates were developed by the addition of 0.04% o-phenylenediamine dihydrochloride (OPD, Sigma-Aldrich Co, Saint Louis, USA) and 0.03% hydrogen peroxide in 0.05 M citrate-phosphate buffer, pH 5.0, and incubated in the dark for 30 min at RT. The reaction was stopped via the addition of 2.5N H2SO4. The absorbances were determined on an automatic microplate ELISA reader (Multiskan plus Labsystem version 2.01) at 492 nm. The cut-off point for serum reactivity was calculated as the mean optical density of the negative reference sera, plus three standard deviations. Serum samples with absorbance values above the cut-off were considered positive. Statistical analysis - The sensitivity, specificity and predictive values for both EITB-IgG and ELISA-IgG tests were calculated using 2 x 2 tables with a confidence interval of 95%. To compare the degree of concordance between the two tests, the kappa index (k) was used. Additionally, the Student's t-test (t) for paired samples at a statistical significance level of 5% was used to assess the effect of serum absorption with A. suum worm extract on both the EITB-IgG and ELISA-IgG tests. RESULTS Using a pool of positive sera from patients with clinical and serological confirmed diagnosis of toxocariasis (four cases of OT and two cases of VLM), a total of 10 antigenic bands were obtained for different batches of TES antigens. Antigenic bands were grouped according to their MW: low MW (LMW: 24, 28, 30 and 35 kDa); intermediate MW (IMW: 48, 56 and 67 kDa) and high MW (HMW: 117, 136 and 152 kDa) (Fig. 1). All serum samples from patients with suspected diagnosis of toxocariasis were positive by the EITB-IgG test (100% of sensitivity), showing the presence of eight major bands (24, 28, 30, 35, 56, 117, 136 and 152 kDa). The 48 and 67 kDa bands were not efficient as the other eight bands for detecting IgG antibodies in these patients. The serum sample from one healthy control reacted with LMW bands where as sera from eight healthy controls reacted with HMW bands (Table I). The sera from patients with other helminth diseases also were analyzed. Sera from 160 and 137 other hel-minth patients tested positive for HMW bands and IMW bands, respectively. Three serum samples from this group also reacted with all the bands. The frequencies of reactivity for each band with sera from different groups are summarized in Table I. When the effect of previous serum absorption with A. suum worm extract was evaluated, LMW and HMW bands were always observed in sera from patients with toxocariasis, while the presence of almost all IMW bands was considerably reduced (Fig. 2). Furthermore, a significant reduction of positivity frequencies for HMW and IMW bands were observed in sera from patients with ascariasis (t = 3.19; p = 0.011), ancylostomiasis (t = 2.86; p = 0.019), enterobiasis (t = 2.93; p = 0.017), hymenolepiasis (t = 2.76; p = 0.022), diphyllobothriasis (t = 2.5; p = 0.034), taeniasis (t = 2.51; p = 0.033) and cysticercosis (t = 2.37; p = 0.042), while sera from patients with trichuriasis, strongyloidiasis, hydatidosis and fascioliasis showed no significant reduction (Table I, Fig. 2). These data suggest that HMW and IMW bands have poor specificities because of their high frequency in patients with other helminth diseases. In contrast, LMW bands (24-35 kDa) were reactive with all sera from patients with toxocariasis, but only with four sera samples from the other groups, yielding 98.6% specificity (95% CI, 96.6-99.5). The sensitivity and specificity for each band of the EITB-IgG test is shown in Table II. The ELISA-IgG test gave positive results for all sera from patients with suspected toxocariasis (100% sensitivity). The same results were found when all of these sera were previously absorbed with A. suum extracts (Table III). Additionally, positive results were also observed in 4.5% and 48.4% of sera from healthy controls and patients with other helminth diseases, respectively. The total specificity of the ELISA-IgG test was 67.9% (95% CI, 62.4-73). Nevertheless, when the same groups of sera were re-tested with pre-absorption with A. suum extracts, the frequency of positivity for sera from patients with other helminth diseases was reduced significantly to 40.32% (t = 3.43; p = 0.005) (Table III). The EITB-IgG test and ELISA-IgG test had a concordance of 100% (k = 1.0; p = 0) in suspected cases of toxocariasis. The concordance of the two tests was 96.3% (k = 0.323; p < 0.001) and 53.7% (k = 0.046; p < 0.01) in healthy controls and patients with other helminth diseases, respectively. The reproducibility of the EITB-IgG test was very good, and there were no existing interferences with sera containing lipids, hemoglobin or billirubin. DISCUSSION The development of specific and sensitive tests to demonstrate the presence of antibodies in sera from patients with suspected toxocariasis is an important step towards improving diagnosis. In this study, we developed and evaluated an EITB-IgG test using TES antigens for the immunodiagnosis of human toxocariasis. The Toxocara larvae secrete a complex set of glycoproteins containing at least 50 distinct components, as enumerated by two-dimensional SDS-PAGE analysis (Page et al. 1991, 1992a), when maintained in vitro and probably in vivo (Maizels et al. 1984, 2000, Maizels & Page 1990, Page et al. 1992a, b). The major constituents of TES antigens are now characterized as mucins, C-type lectins and phosphatidylethanolamine-binding protein and are designated as TES-26, TES-32, TES-45, TES-55, TES-70, TES-120 and TES-400, according to their MWs on SDS-PAGE (Maizels et al. 1984, 2000, Meghji & Maizels 1986, Gems et al. 1995, Gems & Maizels 1996, Loukas et al. 1999, 2000). In our study, we observed bands with a MW pattern that was different than that observed in EITB assays, but we believe that the bands of 24-28 kDa would be closely related with the TES-26, and the bands of 30-35 kDa would be closely related with the major TES-32 band described by Maizels et al. (1984). Sugane and Oshima (1983) have also reported a protein of 35 kDa with a similar MW to one of our antigenic bands within the LMW group. When we analyzed the sera from patients with suspected toxocariasis, we found that the 48 and 67-kDa bands from the IMW group were not recognized as frequently as the other bands, being only recognized in serum samples from some patients with high titers in the ELISA-IgG test (data no shown), thus these bands have poor sensitivity for serodiagnosis of toxocariasis. These antigenic bands might be related to the TES-45, which is secreted in low quantities as reported by Loukas et al. (2000) and Maizels et al. (2000). In another study, Obwaller et al. (1998) reported the presence of a 65-kDa band that binds to IgG antibodies, which might be closely related to our 67-kDa antigenic band. The 56-kDa band from the IMW group might be related to the TES-55 reported by Maizels et al. (2000), but we believe it is more closely related to the 55 kDa band reported by Sommerfelt et al. (2001) in a T. canis experimental infection using a pig model. Iddawella et al. (2007) also reported a specific 57-kDa antigenic band for immunodiagnostics of human toxocariasis. Nevertheless, we have found that this antigenic band had cross-reactions mainly with sera from patients with ascariasis and ancylostomiasis, contradicting their affirmations. The TES antigens show an extensive pattern of bands from 20-400 kDa when analyzed by SDS-PAGE (Maizels et al. 1984, 2000, Speiser & Gottstein 1984). To resolve a wide spectrum of bands, it is necessary to use gradient polyacrylamide gels for separating heterogeneous polypeptide mixtures from high and low MWs, as is the case with the TES antigens. In the present study, we found a characteristic pattern of 10 antigenic bands that were recognized by serum IgG antibodies from patients with clinical and serological confirmed cases of human toxocariasis. The MWs of these components are 24, 28, 30, 35, 48, 56, 67, 117, 136 and 152 kDa. Four of these antigens bands correlate perfectly with LMW bands described by Magnaval et al. (1991). As revealed by western blot, there are two main groups of bands present in the positive cases: one of 24-45 kDa and another group of 66-200 kDa (Magnaval et al. 1991, Nunes et al. 1997, Morales et al. 2002). These studies suggest the HMW bands can cause cross-reactivity and the LMW bands would be more specific for Toxocara. Nunes et al. (1997) identified five major components: above 205 kDa, around 205 kDa, 116-97 kDa, 55-50 kDa and 35-29 kDa. Magnaval et al. (1991) classified the seven antigenic components into two groups, LMW (24-35 kDa) and HMW (132-200 kDa). The number of bands, as well as the exact size of the antigenic bands, varies from one publication to another; the differences can be attributed to the running conditions, the source of MW markers and other multiple inherent factors to the assay, including the presence of enzymatic activity in the TES antigens, which is often not inactivated by enzyme inhibitors added to the antigen solution. In the present study, we have used SDS-denatured and DTT-reduced TES antigens, electrophoretically separated in a 4-16% gradient polyacrylamide gels in order to produce immunoblot strips for the EITB-IgG test. All sera from patients with suspected toxocariasis were positive for the EITB-IgG test, demonstrating 100% sensitivity. However, eight sera from the control group were positive for some bands of the EITB-IgG test, mainly with antigenic bands of the HMW group. These data suggest that these patients were likely infected with other helminthes, such as Enterobius vermicularis, a worm very frequent in Peruvian children (Gilman et al. 1991) or have a mild Toxocara infection. In sera from patients with other helminthiasis, 137 and 160 sera were positive for the IMW and HMW bands, respectively, indicating poor specificities for these bands. Because our country is an endemic area for many helminth infections (except of Schistosoma species) (Gilman et al. 1991, Marcos et al. 2005, Larocque et al. 2006, Romani et al. 2006, Yori et al. 2006, Chero et al. 2007, Cleary et al. 2007, García et al. 2007), we decided to use serum samples from people living in the high Andean Regions, as under these conditions of cold and dry weather, it is unlikely that Toxocara eggs have remained viable (Schantz 1989), thereby reducing the likelihood of co-infection. However, three serum samples from this group were also positive for LMW bands, suggesting concurrent Toxocara infections, since exclusion of a past or recent asymptomatic infection in these individuals was not possible. We have observed a high rate of specificities for antigenic bands from the LMW group described, as the negative control sera did not react with the identified bands (Tables I, II). The ELISA-IgG and EITB-IgG tests showed a 100% of concordance for positive results and 96% of concordance for negative results in healthy controls. However, the ELISA-IgG test has shown positive results in 48.38% of patients with other helminthiasis, indicating poor specificity in regions where the other helminth infections are frequent. The serodiagnosis of human toxocariasis is generally made by ELISA using TES antigens (De Savigny et al. 1979, Jacquier et al. 1991, Roldán et al. 2006). However, because of cross-reactions with other hel-minth diseases, a positive ELISA for Toxocara needs to be confirmed by western blot, which is more specific (Magnaval et al. 2001). In an attempt to improve the specificity of the ELISA-IgG test, many authors have suggested that serum pre-absorption with A. suum antigens can avoid cross-reactivity with other parasites, mainly with other helminths (Camargo et al. 1992, Nunes et al. 1997, Roldán et al. 2006). In a previous report, we found high rates of specificity using this procedure (Roldán et al. 2006). However, we have also observed that this antigenic extract cannot eliminate all cross-reactions, mainly in sera from patients with helminth diseases by cestodes or trematodes. In the present study, we have evaluated the effect of this antigenic extract on the results in both EITB-IgG and ELISA-IgG tests, using a wide range of serum samples. In the group of patients with suspected toxocariasis, the IMW bands were considerably reduced (mainly the bands of 56 and 67 kDa), while the LMW and HMW bands remained positive in these patients. Nunes et al. (1997) have identified one band, with MW around 55-66 kDa, as the band responsible for the cross-reactivity between T. canis and A. suum; this band disappears when previous absorption of serum samples with A. suum antigens is performed. However, Lynch et al. (1988) recognized a band with MW of 81.4 kDa as the band responsible for a strong cross-reactivity between T. canis and A. suum extract. It is possible that reported differences in the MWs of fractions can reflect technical variations in detection and resolution of constituents or differences in methods for maintenance of larvae, such as use of enzymatic inhibitors in these preparations. We have also observed wide variability of results when the serum absorption with A. suum antigens was carried out. We have observed that this antigenic extract could be an effective method to reduce the cross-reactivity when helminth infections by nematodes, such as ascariasis, ancylostomiasis and enterobiasis, are also present. Nevertheless, this antigenic extract cannot reduce the cross-reactivity in nematode infections (such as trichuriasis) and strongyloidiasis. It is possible that the cross-reactive antigens from A. suum are only present in some kinds of nematodes. However, there is also the possibility that the amount of A. suum antigens used in the sera absorption in the present work might not be sufficient to avoid all cross-reactions in these sera. Furthermore, we have found that the A. suum antigens may also be able to reduce cross-reactions in cases of helminth infections by cestodes, such as hymenolepiasis, diphyllobothriasis, taeniasis and cysticercosis, while cases of hydatidosis and fascioliasis are less frequently reduced. Romasanta et al. (2003) have found the existence of cross-reactivity between Fasciola hepatica and T. canis, suggesting that this by taken into consideration when immunoassay tests are used in the diagnosis of parasitic infections. With regard to cysticercosis, García et al. (2007) demonstrated the lack of cross-reactivity with anti-Taenia solium antibodies in the Toxocara-based assays, using an experimental pig model for cysticercosis. We believe that Ascaris extracts would improve the ELISA specificity by reducing the cross-reactivity with some helminth infections, mainly with nematode parasites closely related to A. suum. Nevertheless, it is not a good choice to reduce cross-reactions when cases of strongyloidiasis, hydatidosis or fascioliasis are implicated. Perhaps a possible solution may be the use of a more concentrated antigenic extract to avoid the cross-reactive antibodies from the sera, although Elefant et al. (2006) have used lower concentrations to avoid unspecific antibodies before the ELISA assays. Future studies are also necessary to assess other sources of heterologous antigens that can help to reduce the cross-reactions in the serodiagnosis of human toxocariasis. EITB is a more specific technique than ELISA, but much research concerning the identification of the immunodominant antigens or peptides is still needed. Studies on the purification of specific peptides may obviate the problem of cross-reactivity, and their use at appropriate concentrations can improve the ELISA, a low-cost and easy-to-use method, which appears to be useful for primary screening in endemic areas and a positive result could be confirmed by the EITB procedure. The EITB-IgG test, using the 24-35 kDa, can easily discriminate the true cases of toxocariasis from other helminth infections, without the use of laborious procedures to elevate their specificity in an ELISA test. In conclusion, the EITB-IgG test is a useful tool to confirm the serological diagnosis of human toxocariasis, especially in cases of positive ELISA tests. ACKNOWLEDGEMENTS To Dr. Hector H García, Silvia Rodriguez and Erika Perez (Cysticercosis Working Group in Peru), by the technical assistance in the development of this study, and to Prof. William R Cornejo, by some corrections of this manuscript. REFERENCES
Copyright 2009 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc09065f1.jpg] [oc09065t1.jpg] [oc09065f2.jpg] [oc09065t3.jpg] [oc09065t2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}