 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 104, No. 3, May, 2009, pp. 473-480 ARTICLES Cl gene cluster encoding several small nucleolar RNAs: a comparison amongst trypanosomatids Paola Nocua; Carolina Gómez; Claudia Cuervo; Concepción Puerta+ Laboratorio de
Parasitología Molecular, Departamento de Microbiología, Facultad
de Ciencias, Pontificia Universidad Javeriana, Carrera 7 43-82, Lab. 113, Bogotá,
Colombia P Nocua and C Gómez contributed equally to this work. Financial support: Vicerrectoría Académica, PUJ (1707) Received 8 October
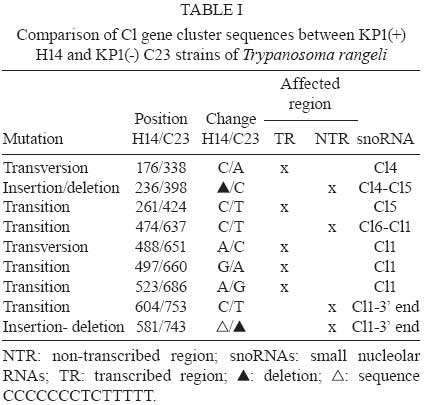
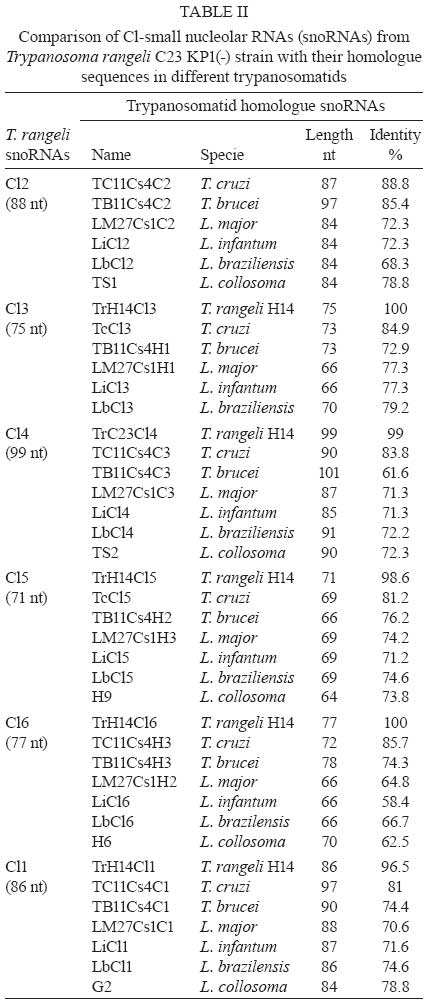
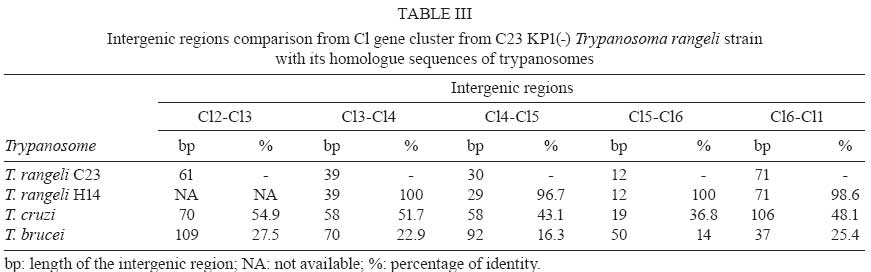
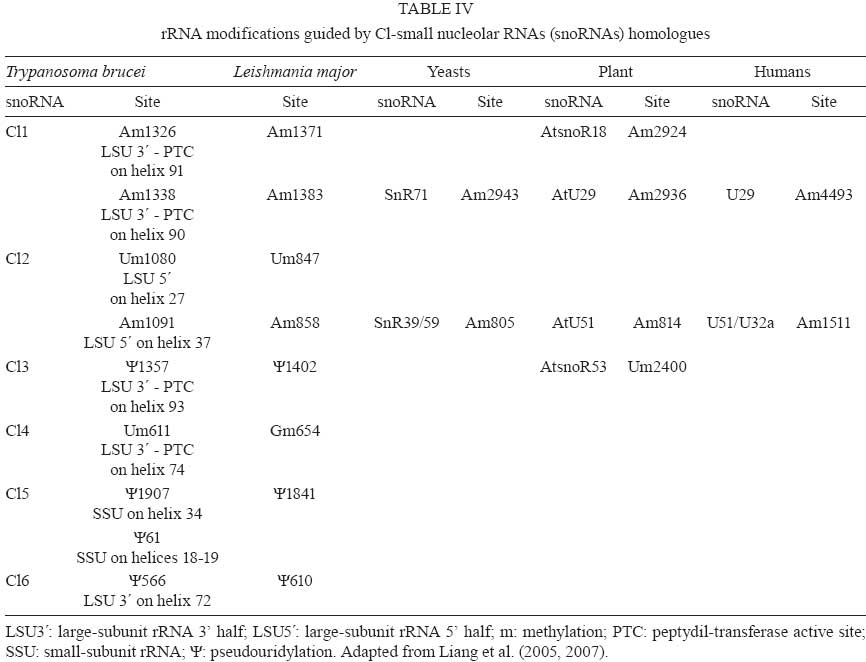
2008 Code Number: oc09075 ABSTRACT Small nucleolar RNAs (snoRNAs) are small non-coding RNAs that modify RNA molecules such as rRNA and snRNA by guiding 2'-O-ribose methylation (C/D box snoRNA family) and pseudouridylation reactions (H/ACA snoRNA family). H/ACA snoRNAs are also involved in trans-splicing in trypanosomatids. The aims of this work were to characterise the Cl gene cluster that encodes several snoRNAs in Trypanosoma rangeli and compare it with clusters from Trypanosoma cruzi, Trypanosoma brucei, Leishmania major, Leishmania infantum, Leishmania braziliensis and Leptomonas collosoma. The T. rangeli Cl gene cluster is an 801 base pair (bp) repeat sequence that encodes three C/D (Cl1, Cl2 and Cl4) and three H/ACA (Cl3, Cl5 and Cl6) snoRNAs. In contrast to T. brucei, the Cl3 and Cl5 homologues have not been annotated in the Leishmania or T. cruzi genome projects (http//:www.genedb.org). Of note, snoRNA transcribed regions have a high degree of sequence identity among all species and share gene synteny. Collectively, these findings suggest that the Cl cluster could constitute an interesting target for therapeutic (gene silencing) or diagnostic intervention strategies (PCR-derived tools). Key words:C/D snoRNA - H/ACA snoRNA - rRNA - Trypanosoma rangeli - trypanosomatid Small nucleolar RNAs (snoRNAs) are a defined population of non-protein coding RNAs that guide post-transcriptional modifications of ribosomal RNA (rRNA) and some spliceosomal small nuclear RNAs (snRNAs) that are crucial for appropriate RNA folding, as well as RNA-RNA and RNA-protein interactions (Weinstein & Steitz 1999). These 60-300 nucleotide (nt)-long RNAs exist as snoRNA-protein complexes called snoRNPs. Most snoRNAs can be divided into two classes on the basis of their function and the presence of conserved sequence motifs; one contains two such motifs, box C (5' -RUGAUGA-3') and box D (5' -CUGA-3') along with two more degenerate internal copies of these elements, C' and D' boxes (Kiss-Laszlo et al. 1998); the other group contains the H (ANANNA) and ACA motifs (Maxwell & Fournier 1995). A few snoRNAs in each family are involved in pre-rRNA processing, but most are associated with nucleotide modification. Box C/D snoRNAs guide 2'-O-ribose methylation (Samarsky et al. 1998), while H/ACA snoRNAs guide pseudouridine formation (Balakin et al. 1996). The snoRNAs interact directly with their RNA targets through base complementarity and methylation or uridine isomerisation reactions are catalysed by core snoRNP proteins. Each box C/D snoRNP contains a single snoRNA harbouring the C and D elements required for snoRNA nucleolar localization, accumulation, maturation and protein association (Samarsky et al. 1998). Box C/D snoRNAs have one or two regions of sequence complementarity to specific region(s) in the rRNA (Bachellerie et al. 1995). These 10-20 nt guide sequences reside immediately upstream of box D or D' and form duplexes with the target RNA, directing methylation of the fifth nucleotide (+5) that is paired upstream of the 5' end of box D or D'; this is known as the "+5 rule" (Decatur & Fournier 2003). The C/D snoRNAs of trypanosomatids such as Leptomonas collosoma (Levitan et al. 1998, Xu et al. 2001, Liang et al. 2004, Uliel et al. 2004), Trypanosoma brucei (Roberts et al. 1998, Dunbar et al. 2000a, b, Liang et al. 2005), Leishmania tarentolae (Roberts et al. 1998), Leishmania major (Uliel et al. 2004, Liang et al. 2007), Trypanosoma cruzi (Roberts et al. 1998, Uliel et al. 2004) and Trypanosoma rangeli (Morales et al. 2002) exhibit the same general structure and follow the +5 rule as well. Some trypanosomatid C/D snoRNAs are also able to guide methylation at two different sites in the same rRNA molecule or even in two different rRNAs, acting as double guides (Morales et al. 2002). Box H/ACA snoRNAs have two conserved elements: the H domain (located in a hinge region joining two functionally similar hairpin domains) and the ACA triplet located 3 nt upstream from the 3' end. Two short rRNA recognition motifs within the snoRNA pair with rRNA sequences flanking the target uridine, which is always 14-16 nt upstream of the snoRNA H or ACA box (Ganot et al. 1997). The two internal loops within the H/ACA snoRNA usually contain complementary rRNA domains (Ni et al. 1997). These snoRNAs form a single hairpin in trypanosomatids ending in the AGA triplet instead of ACA (Liang et al. 2001, 2002, 2004, 2007, Barth et al. 2005). Arranged differently from mammalians, most of trypanosomatid snoRNAs are clustered in tandem repeats and carry a mix of both snoRNA families (Dunbar et al. 2000a, Morales et al. 2002, Liang et al. 2007). This genomic organisation resembles that of plants in which snoRNAs are also clustered and transcribed as polycistronic snoRNA precursors (Brown et al. 2003). T. rangeli is a hemoflagellate protozoan parasite that, in contrast to T. cruzi (the etiologic agent of Chagas disease), is considered to be non-pathogenic to mammalian hosts (D'Alessandro & Saravia 1999). Despite its lack of human pathogenicity, T. rangeli is a serious concern for human Chagas disease epidemiology and diagnosis (Guhl & Vallejo 2003). Recent studies have classified T. rangeli into KP1(+) and KP1(-) strains, which are related to transmission-vector ability (Vallejo et al. 2003, 2007). The Cl gene cluster (coding for several snoRNAs) was characterised in this work in a representative strain from each T. rangeli group and compared to those from T. cruzi, T. brucei, L. major, Leishmania infantum, Leishmania braziliensis and L. collosoma. MATERIALS AND METHODS Parasites -Epimastigotes from the T. rangeli KP1(+) H14 strain (MHOM/Hond/H14) (Acosta et al. 1991) were used in this study. They were characterised by using S35/S36/KP1L PCR (Vallejo et al. 2002). Parasites were grown at 26ºC in modified LIT medium supplemented with 15% (v/v) heat-inactivated foetal bovine serum. PCR amplification, cloning and nucleotide sequence - PCR was performed using TrF1 (5' - CGC CCC GTC TTG CCC TGT-3') and TrR2 (5' - CGC AGC AAG GAC AGG AGG GA-3') primers, which, based on the T. rangeli KP1(-) C23 (MAOT/CO/82/C23) (Zuñiga et al. 1997) strain, amplify a 620 bp fragment exclusively in T. rangeli. A 25 µL reaction contained 100 ng purified genomic DNA, 1X reaction buffer (10 mM Tris-HCl pH 9.0, 50 mM KCl and 0.1% Triton X-100), 1.25 units of Expand High Fidelity enzyme (Roche, Branford, USA), 1.5 mM MgCl2, 0.2 mM of each deoxynucleoside triphosphate and 20 pmol of each primer. Reactions were carried out in an MJ Research PTC-100 DNA thermal cycler using the following method: 94ºC /5 min, 15 cycles of 95ºC/30 s, 63ºC/1 min, 72ºC/30 s and 20 cycles of 95ºC/30 s, 60ºC/1 min, 72ºC/30 s and a final incubation of 72ºC for 5 min. Amplified fragments were purified from an agarose gel using a GFX Gel Band Purification kit (Amersham Biosciences) and cloned into the pGEM-T Easy plasmid (Promega). Both cloned fragment strands were sequenced by the Sanger method (Sanger et al. 1977) in a 373 Automatic DNA sequencer (Pharmacia LKB) using universal and specific primers. Sequence analysis - Sequence of an 801 bp fragment from the T. rangeli KP1(-) C23 strain was retrieved from GenBank (accession AY028385) and included for comparative analyses. The T. rangeli KP1(+) H14 strain sequence produced in this study is available from the GenBank database (accession EF100612). Homology searches were performed in GenBank and parasite genome project databases (http://www.genedb.org) using the BLAST program (Stephen et al. 1997); pair-wise and multiple sequence alignments were carried out using LALIGN (Pearson 1990) and ClustalW (Thompson et al. 1994) software, respectively. RESULTS Analysing the Cl gene cluster from T. rangeli - BLASTN comparative analysis of the previously reported 801 bp fragment coding for snoRNA-Cl1 of the T. rangeli C23 strain (Morales et al. 2002) and T. brucei GeneDB version 4.0 revealed the presence of additional genes coding for five snoRNAs. This gene cluster repeat (named Cl) encoded six snoRNAs, three C/D (Cl1, Cl2 and Cl4) and three H/ACA (Cl3, Cl5 and Cl6), organised from the 5' end from Cl2, Cl3, Cl4, Cl5, Cl6 to Cl1. All C/D snoRNAs exhibited the consensus C, D, C' and D' boxes and regions having sequence complementarity to the methylation site on rRNA (Fig.1). Like other trypanosomatids, Cl gene cluster H/ACA snoRNAs formed single hairpins ending in the AGA motif (Fig.2). Amplified sequence from the Cl gene cluster in the T. rangeli H14 strain (corresponding to nts 163-782 of the C23 strain) revealed 96.5% identity between the sequences of both strain, exhibiting two transversions, five transitions and two insertion-deletions (Table I). Comparing the T. rangeli Cl gene cluster sequence with its homologue in T. cruzi - Comparative analysis of the T. rangeli C23 sequence with the T. cruzi genome revealed 81-88% identity between Cl1 and TC11C4C1, Cl2 and TC11C4C2, Cl4 and TC11C4C3 and Cl6 and TC11C4H3 (Table II). Since the current T. cruzi genome assembly and annotation version is fragmented and redundant, all contigs containing the Cl gene cluster were aligned with T. rangeli sequences. The analysis revealed the presence of Cl3 and Cl5 homologues in T. cruzi (84.9% and 81.2% identity, respectively). It was also found that the gene order within the Cl cluster was conserved in both trypanosomes. In fact, transcribed regions were highly conserved between both species whereas non-transcribed regions differed in both size and sequence (Table III). The genomic sequence contained 103 contigs that include Cl-snoRNAs (T. cruzi GeneDB, version 4.0). Some of them, like contig 7066, carried other genes such as those coding for phosphatidylinositol kinase (Tc00.1047053506719.10), which corresponds to orthologues Tb11.47.0002 and LmjF27.0890 in T. brucei and L. major, respectively. Comparing the T. rangeli Cl gene cluster sequence with its homologue in other trypanosomatids - Homologues for all Cl genes were detected in T. brucei, L. major, L. infantum and L. braziliensis, having 61.6-85.4% identity with T. brucei and 58.4-79.2% with the Leishmania species (Table II). All Cl-snoRNAs of L. collosoma were found to share 62.5-78.8% identity with those T. rangeli, except for Cl3 (Table II). Multiple alignments of all trypanosomatid snoRNAs revealed conservation of both sequence and position for the characteristic motifs of each snoRNA family (Figs 1, 2). Table IV shows the rRNA modifications carried out by Cl-snoRNA homologues from trypanosomatids, yeasts, plants and humans. Cl gene cluster organisation and genome location was studied in the trypanosomatid genome projects. This cluster in T. brucei was repeated five times, consisting of 32 snoRNA genes that maintained the same T. rangeli snoRNA gene order and were located on the antisense strand of chromosome 11 in position 456973-461605 (T. brucei GeneDB, version 4.0). This cluster was flanked upstream by a gene encoding the isovaleryl-CoA dehydrogenase protein (Tb11.55.0026) and downstream by two ESAG genes (Tb11.55.0027 and Tb11.55.0028). It is worth noting that a phosphatidylinositol kinase gene (Tb11.47.0002) was located proximally on the sense strand (position 423851-428194). The Cl gene cluster in L. major is located on both strands of chromosome 27. Two clusters of Cl2, Cl4, Cl5 and Cl1 are located in the sense strand at positions 369619- 370773, preceded upstream by a phosphatidylinositol kinase (LmjF27.0890) and a hypothetical conserved protein (LmjF27.0900) and followed downstream by another hypothetical conserved protein (LmjF27.0910). Fifty three snoRNAs are organised in the antisense strand into two arrays (Cl1, Cl3 and Cl6) and (Cl1, Cl2, Cl4 and Cl5), which are intercalated and repeat five and nine times, respectively. They are located at positions 375179-383161, flanked upstream by a hypothetical conserved protein (LmjF27.0920) and the isovaleryl-CoA dehydrogenase protein (LmjF27.0930). Cl gene cluster snoRNAs have not been annotated yet in the L. infantum and L. braziliensis genome projects. However, BLASTN analysis has revealed their presence on chromosome 27 from both species. Forty six snoRNAs, organised similarly to those from the antisense strand of L. major, were located in the antisense strand in L. infantum at position 310106-316427 (L. infantum GeneDB, version 3.0). They were flanked upstream by the isovaleryl-CoA dehydrogenase protein (LinJ27_V3.0790) and a hypothetical conserved protein (LinJ27_V3.0780) and followed downstream by another hypothetical conserved protein (LinJ27_V3.0770). A phosphatidylinositol kinase gene (LinJ27_V3.0750) was located proximally on the sense strand (position 295857-301364). In the case of L. braziliensis, there were 22 snoRNAs organised into two arrays (Cl2, Cl4, Cl5 and Cl1) and (Cl3, Cl6 and a truncated Cl1), which are intercalated and repeat four and two times, respectively. This cluster was located in the sense strand in position 380881-384367 (L. braziliensis GeneDB, version 2.0) flanked upstream by a phosphatidylinositol kinase protein (LbrM27_V2.0970) and a hypothetical conserved protein (LbrM27_V2.0980) and followed downstream by another hypothetical conserved protein (LbrM27_V2.0990). An isovaleryl-CoA dehydrogenase protein (LbrM27_V2.1010) was located proximally in the antisense strand (positions 393327-394559). Interestingly, two snoRNAs (Cl2 and Cl1) were found 4276 nt downstream of the Cl gene cluster in position 388642-388849. It is worth noting that the copy number of each Cl-snoRNA varied within and among species, being more abundant in L. major than in the other species (Table V). DISCUSSION Non-protein coding RNAs (ncRNA) play critical roles in different processes affecting protein synthesis. snoRNAs (a type of ncRNA) are involved in RNA modifications leading to correct RNA folding and RNA-RNA and RNA-protein interactions (Zemann et al. 2006). It is known that snoRNAs from trypanosomatids are involved not only in rRNA processing and modifications but also in snRNA modifications thereby affecting protein synthesis and trans-splicing (Barth et al. 2005, 2008). In this study, the Cl gene cluster encoding several snoRNAs was characterised in T. rangeli and compared to those from T. cruzi, T. brucei, L. major, L. infantum, L. braziliensis and L. collosoma. In accordance with their evolutionary origin, trypanosomatid Cl-snoRNAs present particular features shared with those from euglenids: H/ACA snoRNAs consist of a single hairpin (Russell et al. 2004), boxes C' and D' from C/D snoRNAs can be easily distinguished in spite of exhibiting some variations (Russell et al. 2006) and functional isoforms allow the presence of some plasticity in the 5' half of transcribed regions (Liang et al. 2005, Russell et al. 2006). Consistent with previous reports showing that snoRNAs in trypanosomatids are encoded by clusters of arrayed tandem genes, the Cl gene cluster repeat contains six snoRNA genes exhibiting an intercalated array of C/D and H/ACA snoRNAs. Given the fact that the parasite requires large amounts of mature rRNA molecules, the arrangement of several alternating snoRNA genes might provide a solution to a lack of transcriptional controls in these parasites. The Cl gene cluster order, size and sequence is highly conserved between the KP1(+) and KP1(-) strains from T. rangeli, having few mutations that affect both the transcribed and non-transcribed regions. The biological significance of this finding needs to be addressed by analysing more strains. Nevertheless, it is reasonable to assume that the observed changes, especially transversion, could affect the spatial structure and function of the RNA molecule. We observed that transcribed regions and gene order of the Cl gene cluster were conserved among the three species of trypanosomes studied. However, Cl-snoRNA and intergenic spacer sequence identity was higher with T. cruzi homologues than with those of T. brucei; this finding agrees with previous reports revealing a closer T. rangeli and T. cruzi phylogenetic relationship than that between T. rangeli and T. brucei (Maia da Silva et al. 2004, Diez et al. 2005, Cuervo et al. 2006). Orthologues from T. cruzi phosphatidylinositol kinase (Tc00.1047053506719.10), Tb11.47.0002 and LmjF27.0890, surround the Cl-snoRNA array in T. brucei and L. major. Consequently, it is possible that the T. cruzi Cl gene cluster is located in the gene synteny region similar to what has been reported for chromosomes 11 and 27 of T. brucei and L. major, respectively (El-Sayed et al. 2005). Interestingly, Cl gene cluster members of leishmanias were found in different tandem arrays from those observed in trypanosomes. Taking into account the efficient expression of trypanosomatid snoRNAs, one could hypothesize that expression is influenced by extended stems formed in the extragenic flanking sequences of adjacent snoRNA molecules (Liang et al. 2007); these array order differences could have important biological consequences. T. rangeli intergenic regions of the Cl gene cluster range from 12-93 nt. This range is in accordance with the minimum 10 nt distance needed for the proper processing of each snoRNA (Xu et al. 2001, Liang et al. 2004). Indeed, Cl gene cluster C/D box snoRNA expression has been described in L. collosoma (Xu et al. 2001) and T. brucei (Barth et al. 2008). Although there is no specific expression data from Cl-H/ACA snoRNAs, other B2 cluster H/ACA snoRNAs have been detected in L. collosoma by Northern blot or primer extension analysis (Xu et al. 2001, Liang et al. 2004) suggesting the expression of these snoRNAs as well. An intense signal corresponding to approximately 90 nt has been shown in Northern blot assays in T. rangeli using the total Cl cluster as a probe, which seems to include all Cl-snoRNAs (Morales et al. 2002). In spite of having the same copy number, C/D snoRNAs in T. brucei have different levels of expression, with Cl1 showing the greatest. However, snoRNAs Cl2 and Cl3 exhibited greater target methylation as compared to Cl1 (Barth et al. 2008). These results showed that other factors aside from copy number and expression level were influencing the modification guiding process, such as the presence of secondary structure at the modification site. The Cl-snoRNA copy numbers from leishmanias differ among each snoRNA according to its array. In contrast to C/D snoRNAs Cl1 and Cl2, H/ACA snoRNAs Cl3 and Cl6 are less represented in the genome. This dosage effect might be a compensatory mechanism driven by a need for Cl-snoRNA differential expression. Likewise, in archaea and plants some Cl-snoRNAs can function as double guides, guiding two modifications at proximal (Cl1 and Cl2) and distal (Cl5) sites on the same rRNA molecule (Omer et al. 2000, Brown et al. 2003). It was especially interesting that all Cl-snoRNA sequences were highly conserved amongst all trypanosomatids, ranging from 61.6-88.8% identity for C/D and from 58.4-85.7% for H/ACA snoRNAs. This finding suggests it is important for these parasites to maintain the rRNA modifications performed by Cl-snoRNAs. Methylation and pseudouridylation mapping carried out by Cl-snoRNA homologues in T. brucei and L. major has revealed that four modifications (Am1326, Am1338, Um611 and ¨1357) are located within the LSU rRNA peptidyl transferase active site. Although blocking individual rRNA modifications has not had any effect, King et al. (2003) have shown that depleting multiple modifications in the LSU reaction centre region have had synergistic, negative effects on growth. It has been reported recently that position U611 within the LSU is 66.2% hypermethylated in bloodstream versus procyclic forms; this may help the parasite to adapt to a higher vertebrate host temperature (Barth et al. 2008). It is of the upmost importance that homologues that carry out Am1338 (Cl1) and Am1091 (Cl2) LSU modifications have been found in yeast and even humans, implying an importance of these modifications for rRNA structure and function. On the other hand, other Cl-snoRNA modifications such as Um1080 (Cl2), Um611 (Cl4), Ψ 1907, Ψ 61 (Cl5), and Ψ566 (Cl6) seem to be trypanosomatid-specific. The finding that trypanosomatid Cl-snoRNAs share important traits such as sequence, function and specificity renders this cluster a good target candidate for medically important interventions of these parasites. For instance, a therapeutic gene expression silencing approach could be addressed. In fact, Liang et al. (2003) have reported the silencing of T. brucei TBC4 C/D snoRNA. On the other hand, species-specific differences in the non-transcribed regions of Cl-snoRNAs could be useful for developing PCR-based diagnostic tools. Indeed, Morales et al. (2002) have developed a PCR test specific for T. rangeli detection, which does not amplify the DNA of any T. cruzi groups (Pavia et al. 2007). ACKNOWLEDGEMENT To Dr. RS Nicholls and M Montilla, from the Parasitology Laboratory, Instituto Nacional de Salud, for kindly providing the Trypanosoma rangeli H14 strain. REFERENCES
Copyright 2009 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc09075t1.jpg] [oc09075f1.jpg] [oc09075t2.jpg] [oc09075t4.jpg] [oc09075t5.jpg] [oc09075t3.jpg] [oc09075f2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}