 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 104, No. supp 1, July, 2009, pp. 270-274 α-N-acetylglucosamine-linked O-glycans of sialoglycoproteins (Tc-mucins) from Trypanosoma cruzi Colombiana strain Adriane R TodeschiniI; Eliane G de AlmeidaI; Orlando A AgrellosI; Christopher JonesII; José O PreviatoI; Lucia Mendonça-PreviatoI, + IInstituto
de Biofísica Carlos Chagas Filho, Centro de Ciências da Saúde,
Universidade Federal do Rio de Janeiro, 21949-900 Rio de Janeiro, RJ, Brasil Received 12 May
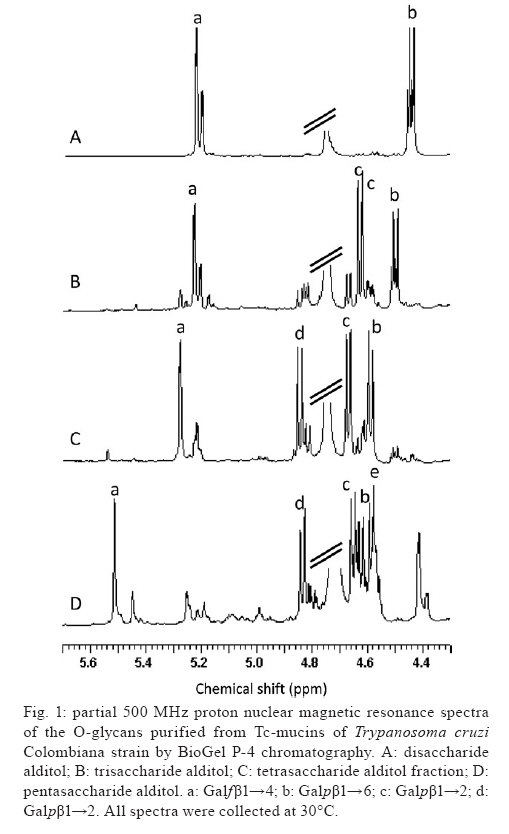
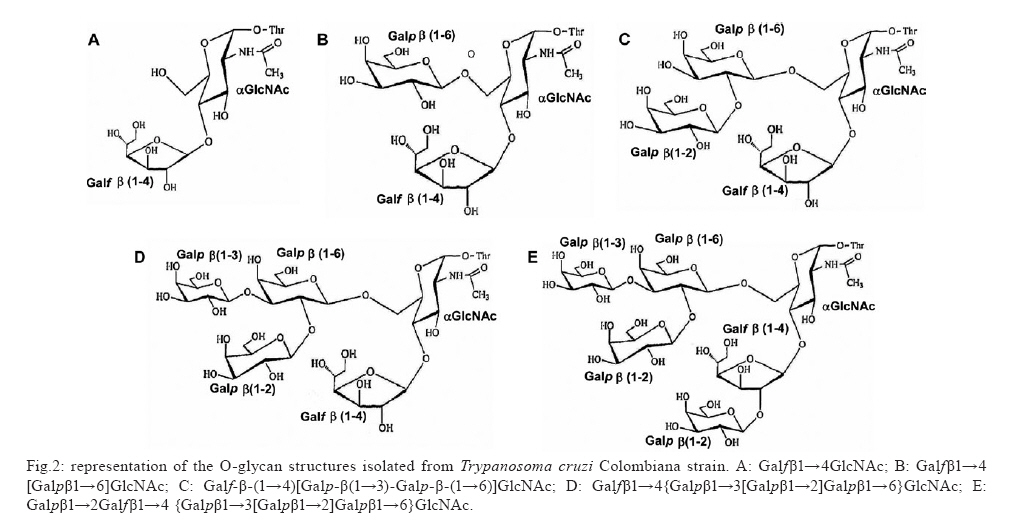
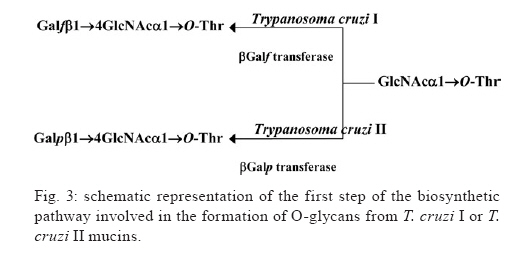
2009 Financial support: CNPq, FAPERJ, National Institute for Science and Technology in Vaccines ART and EGA contributed equally to this study. Code Number: oc09117 ABSTRACT Trypanosoma cruzi sialoglycoproteins (Tc-mucins) are mucin-like molecules linked to a parasite membrane via a glycosylphosphatidylinositol anchor. We previously determined the structures of Tc-mucin O-glycan domains from several T. cruzi strains and observed significant differences among them. We now report the amino acid content and structure of Tc-mucin O-glycan chains from T. cruzi Colombiana, a strain resistant to common trypanocidal drugs. Amino acid analysis demonstrated the predominance of threonine residues (42%) and helped to identify the O-glycans as belonging to a Tc-mucin family that contain a β-galactofuranose (β-Galf) residue attached to an α-N-acetylglucosamine (α-GlcNAc) O-4, with the most complex glycan, a pentasaccharide-GlcNAc-ol with a branched trigalactopyranose chain, on the GlcNAc O-6. The presence of β-Galf on O-glycans from T. cruzi Colombiana mucins supports the use of glycosylation as a phylogenetic marker for the classification of Colombiana in the T. cruzi I group. Key words: Trypanosoma cruzi - NMR spectroscopy - glycoconjugate - Colombiana strain - mucins - sialic acid Chagas disease, caused by the protozoan parasite Trypanosoma cruzi, affects nearly eight million people in South and Central America (WHO 2007) and is becoming a significant health problem in developed countries such as the United States and Spain (Riera et al. 2006, Hotez 2008). The infective forms of T. cruzi, known as trypomastigotes, invade mammalian cells by a not yet completely defined mechanism. However, glycoconjugates on the parasite cell surface have been implicated in adhesion to and penetration of host cells and in modulation of the immune system of the mammalian host (Travassos & Almeida 1993, Almeida & Gazzinelli 2001, Previato et al. 2004, Buscaglia et al. 2006). Among the most abundant glycoproteins on the T. cruzi cell surface are Tc-mucins, trans-sialidase (TS) family members and the mucin-associated surface proteins (MASPs) (Bartholomeu et al. 2009,). It is estimated that there are about 850 mucin-like genes, which, when grouped with the TSs, TS-like genes and MASPs, correspond to about 7% of the T. cruzi genome (El-Sayed et al. 2005). The Tc-mucins are highly O-glycosylated, threonine-rich glycoproteins (Almeida et al. 1994, Previato et al. 1994, 1995, Acosta-Serrano et al. 1995, Di Noia et al. 1996, Mendonça-Previato et al. 2008) that are glycosylphosphatidylinositol-anchored to the parasite surface (Previato et al. 1995). O-linked oligosaccharides account for up to 60% of the mucin molecular mass (Previato et al. 1994) and are linked to Thr residues through an N-acetylglucosamine (α-GlcNAc) unit, which is added by an unusual α-O-N-acetylglucosaminosyl transferase (Previato et al. 1998). The parasite mucins incorporate α2,3-linked sialic acid from exogenous sialyl-glycoconjugates (Previato et al. 1985, 1990) by means of a glycoside hydrolase TS (Previato et al. 1985). Sialylation of the T. cruzi surface by the enzyme TS is pivotal for the establishment of infection (Frasch 2000). Up to the present time, the O-glycan chains of T. cruzi mucins have been characterised from the epimastigote of five strains. O-glycan chains are composed of GlcNAc, galactose (Gal) and sialic acid (Neu5Ac); however, they can vary in complexity and in the ring form of the galactosyl residue in a strain-dependent manner (Previato et al. 1994, 1995, Todeschini et al. 2001, Agrellos et al. 2003, Jones et al. 2004). It has been verified that whilst O-glycans from the mucins of Y (Previato et al. 1995) and CL Brener (Todeschini et al. 2001) strains do not contain β-Galf residues as a substituent on the α-GlcNAc O-4, this substituent is observed in G, Dm28c and Tulahuen strains (Previato et al. 1994, Agrellos et al. 2003, Jones et al. 2004). Several laboratories have reported the division of T. cruzi into two major groups with distinct designations. To homogenise the nomenclature, the groups were named T. cruzi I and T. cruzi II by consensus (Anonymous 1999). Interestingly, Tc-mucin O-glycan core structures correlate with the classification of T. cruzi strains based on the size of the PCR products of the 24S alpha rRNA and the mini-exon genes (Souto et al. 1996). Strains whose mucins contain Galf (G, Dm28c and Tulahuen) group together in T. cruzi I and the strains lacking Galf are classified as T. cruzi II (Jones et al. 2004). The Colombiana strain, isolated from a chronic human case in Colombia (Federici et al. 1964), is particularly interesting since it is resistant to drugs used to treat Chagas disease (Murta et al. 1998). Genetic analysis based on the dimorphism of the 24S alpha rRNA gene and the intergenic region of the mini-exon gene (Souto et al. 1996) indicated that the Colombiana strain belongs to the T. cruzi I group. This work reports a structural study of the mucins from epimastigote forms of T. cruzi Colombiana strain and emphasises the potential use of structural data as a taxonomic parameter. MATERIALS AND METHODS T. cruzi growth - Epimastigote forms of T. cruzi Colombiana strain were cultivated in brain heart infusion medium supplemented with 10 mg/L hemin, 20 mg/L folic acid and 5% (v/v) foetal calf serum at 28ºC under constant agitation. Cultures were harvested during the exponential phase of growth and frozen as previously described (Previato et al. 1994). Isolation and purification of Tc-mucin - Frozen cells (50 g wet weight) were thawed and extracted twice with cold water (150 mL) and the pellet was recovered by centrifugation (12,000 x g, 10 min, 4°C). The pellet was extracted with 45% (v/v) aqueous phenol at 75°C for 15 min. The aqueous phase was dialysed against tap water, lyophilised, dissolved in water and applied to a 4.7 x 25 cm column of Bio-Gel P-60. Excluded material containing carbohydrate was freeze-dried and the mucin was recovered in the insoluble phase after extraction (twice) with chloroform/methanol/water (10:10:3, v/v/v) (Previato et al. 1994). Water-soluble material in the chloroform/methanol/water insoluble components was recovered and lyophilised. The homogeneity of the mucin was determined by electrophoresis in a 15% polyacrylamide gel in the presence of sodium dodecyl sulphate (SDS-PAGE) (Laemmli 1970) and the bands were visualised by staining with periodate-Schiff (PAS) reagent (Fairbanks et al. 1971). Isolation and fractionation of O-glycan-alditols -Tc-mucin was submitted to base-catalysed β-elimination with concomitant reduction using 20 mM NaOH and 0.3 M NaBH4 at 37°C for 24 h. The solution was then neutralised with 50% (v/v) acetic acid, passed through a Dowex 50W-X8 ion-exchange column and lyophilised (Previato et al. 1994). The material was dissolved in water and fractionated on a Bio-Gel P-4 column (1 x 110 cm). Fractions of 1.25 mL were collected and the carbohydrate elution was monitored by spotting 4-μL portions onto a thin layer chromatography plate and staining with orcinol-H2SO4 spray (Humbel & Collart 1975). Carbohydrate-containing fractions were further fractionated by high performance liquid chromatography on porous graphitic carbon as previously described (Jones et al. 2004). Carbohydrate analysis - The neutral sugar composition was determined after methanolysis in 0.5 M methanolic HCl as previously described (Previato et al. 1994). Amino acid analysis - Amino acid analysis was determined after acid hydrolysis in 5.6 N HCl at 110ºC for 22 h in an autoanalyser by the method of Fauconnet and Rochemont (1978). Nuclear magnetic resonance spectroscopic (NMR) analysis - Tc-mucin O-glycan samples were analysed by NMR on a Varian Unity 500 spectrometer as previously described (Previato et al. 1994). RESULTS Isolation of Tc-mucin, amino acid and carbohydrate analysis - Mucin from epimastigote forms of T. cruzi Colombiana strain was extracted with aqueous phenol and recovered from the water-soluble fraction of the chloroform/methanol/water insoluble components. Analysis of this material by SDS-PAGE revealed diffuse PAS staining of bands with apparent molecular masses of 45 and 43 kDa. Carbohydrate analysis of the Tc-mucins indicated the presence of Gal, GlcNAc and mannose (Man) in a molar ratio of 4.0:1.3:1.0 along with Neu5Ac. Amino acid analysis demonstrated Thr to be the predominant amino acid, contributing 42% of the total, whilst Asp, Glu, Ala, Pro contributed 12.5%, 12.6%, 8.8%, 8.1% and 7.2%, respectively. All other remaining amino acids comprised 8.3% of the total. O-linked glycans from Tc-mucins - O-glycans were released from mucins by reductive β-elimination and fractionated on a BioGel P4 column. The resulting six carbohydrate-containing fractions were analysed by NMR spectroscopy. The O-glycan alditols were identified by comparing their one-dimensional 1H spectra (Fig. 1) with those of previously characterised O-glycans (Mendonça-Previato et al. 2008). All of the fractions also contained a ManNAc-ol analogue, a product of the base-catalysed epimerisation of GlcNAc (Jones et al. 2000), as was evident from a chemical shift of the N-acetyl methyl resonance at 2.03 ppm for ManNAc-ol and 2.05 ppm for the GlcNAc epimers (data not shown). The lowest molecular mass material was identified as GlcNAc-ol, whilst the monosaccharide alditol fraction contained a single major component identified as Galfβ1→4GlcNAc-ol (data not shown). This was evidenced by a single intense anomeric signal at a lowfield position (5.154 ppm) with a small coupling constant (3JH1,H2 = 2.3 Hz) characteristic of a β-Galf residue. This compound was previously characterised in the reductive cleavage products of the mucins from T. cruzi G-strain (Acosta-Serrano et al. 1995), the Dm28c clone (Agrellos et al. 2003) and the Tulahuen strain (Jones et al. 2004). The disaccharide alditol fraction contained two components present in an approximate ratio of 6:4 (Fig. 1A). The minor component was Galpβ1→2Galfβ1→4ManNAc-ol, which had been observed in a study of the Tulahuen strain (Jones et al. 2004). The major component contains the presence of an anomeric signal at 4.45 ppm, typical of a terminal β-Galp residue. Disubstitution of the GlcNAc-ol at O-4 and O-6 was determined by comparison of the 1D 1H NMR spectrum with that of the disaccharide alditol previously isolated from G (Previato et al. 1994) and Tulahuen strain mucins (Jones et al. 2004) and is consistent with the Galfβ1→4 [Galpβ1→6]GlcNAc-ol structure. Compared with the disaccharide alditol, the one-dimensional 1H NMR spectrum of the trisaccharide alditol fraction (Fig. 1B) contained an additional anomeric signal at 4.62 ppm, characteristic of a terminal Galpβ1→3 residue, and a shift of the Galpβ1→6 H-1 resonance from 4.45 ppm to 4.50 ppm. This component was therefore identified as the known compound Galf-β-(1→4)[Galp-β(1→3)-Galp-β-(1→6)]GlcNAc-ol (Previato et al. 1994, Jones et al. 2004). The 1D 1H spectra of the tetrasaccharide alditol fraction (Fig. 1C) showed the presence of a single β-Galf spin system and three β-Galp spin systems at chemical shifts identical to those previously reported for the structure of Galfβ1→4{Galpβ1→3[Galp β1→2]Galpβ1→6}GlcNAc-ol in studies of the O-glycans from T. cruzi G (Previato et al. 1994) and Tulahuen strain mucins (Jones et al. 2004). The most complex structure isolated was a pentasaccharide alditol. The 1H NMR spectra of this fraction (Fig. 1D) showed an additional β-Galp spin system and a downfield shift of the β-Galf H-1 and H-2 from 5.3 - 5.5 ppm (β-Galf H-1) and from 4.08 - 4.41 ppm (H-2). These data, and the TOCSY spectrum (datum not shown), are identical with those previously reported for the structure of Galpβ1→2Galfβ1→4 {Galpβ1→3[Galpβ1→2]Galp β1→6}GlcNAc-ol from the G-strain (Previato et al. 1994). DISCUSSION T. cruzi infects a broad range of hosts including wild and domestic animals. In humans, it causes Chagas disease and has a variety of clinical presentations. The severity and symptoms of the disease vary with the geographical region and are related to the characteristics of the patient and the parasite. T. cruzi strains diverge in many other aspects, including tissue tropism, morphology, infectivity, isoenzymes and drug resistance. Early studies provided evidence that T. cruzi is diploid, that sexual reproduction is rare or absent and that the population evolved from independent clonal lineages (Tibayrenc et al. 1986, Tibayrenc & Ayala 1988). Indeed, the remarkable genetic diversity observed raises the question of whether all of these strains are part of a single species or include more than one (Macedo & Pena 1998). Together, these observations require a means to group the strains and a number of classifications have resulted. Molecular typing of T. cruzi strains based on the size of the PCR products of the 24S alpha rRNA and the mini-exon genes (Souto et al. 1996) indicated that the G, Dm28c and Tulahuen strains belong to the T. cruzi I Group and that the Y and CL Brener strains belong to the T. cruzi II Group. Nevertheless, it should be emphasised that CL Brener, the reference strain of the T. cruzi genome project, is a hybrid (El Sayed et al. 2005). Great efforts have been made to understand this heterogeneity. Biological and epidemiological studies provide support for an association of T. cruzi Group II with the domestic cycle, which is mostly involved in human infections, while T. cruzi Group I is associated with the sylvatic cycle, which is asymptomatic in humans most of the time (Briones et al. 1999). The observed variability during infection by different T. cruzi strains is most probably a result of the diversity in parasite/host interactions resulting from variability of the molecules expressed on both the parasite and host cell surfaces. Mucins are among the most abundant glycoproteins on the T. cruzi surface. They are Thr-rich, highly O-glycosylated glycoproteins. The Colombiana strain mucins appear as a double band of 43/45 kDa by SDS-PAGE. The O-glycan chains (Fig. 2) coincide with the Tc-mucins described on strains classified as belonging to T. cruzi group I (Anonymous 1999) (Tulahuen, Dm28c, G and Tulahuen) and contain Galfβ1→4 substituents (Previato et al. 1994, Agrellos et al. 2003, Jones et al. 2004), which have not been observed in T. cruzi group II (Anonymous 1999) strains (Y and CL Brener) (Previato et al. 1995, Todeschini et al. 2001). These observations may be of epidemiologic significance since mammalian hosts do not produce glycoconjugates containing Galf and present a strong immune response against this epitope (Mendonça-Previato et al. 1983, Golgher et al. 1993). Together, these data suggest that structural biology data can be used as an approach to classify/identify T. cruzi strains. Herein we emphasise the importance of accumulating structural data for such comparisons and propose the use of these data as a taxonomic parameter in the differentiation of the T. cruzi clones (Fig. 3). REFERENCES
Copyright 2009 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc09117f3.jpg] [oc09117f2.jpg] [oc09117f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}