 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 104, No. supp 1, July, 2009, pp. 281-287 Swimming against the current: genetic vaccination against Trypanosoma cruzi infection in mice Mauricio M RodriguesI, +; Bruna C de AlencarI; Carla ClaserI; Fanny TzelepisI; Eduardo L SilveiraI; Filipe A HaollaI; Mariana R DominguezI; José Ronnie VasconcelosI, II, III ICentro
Interdisciplinar de Terapia Gênica (CINTERGEN) Received 10 March
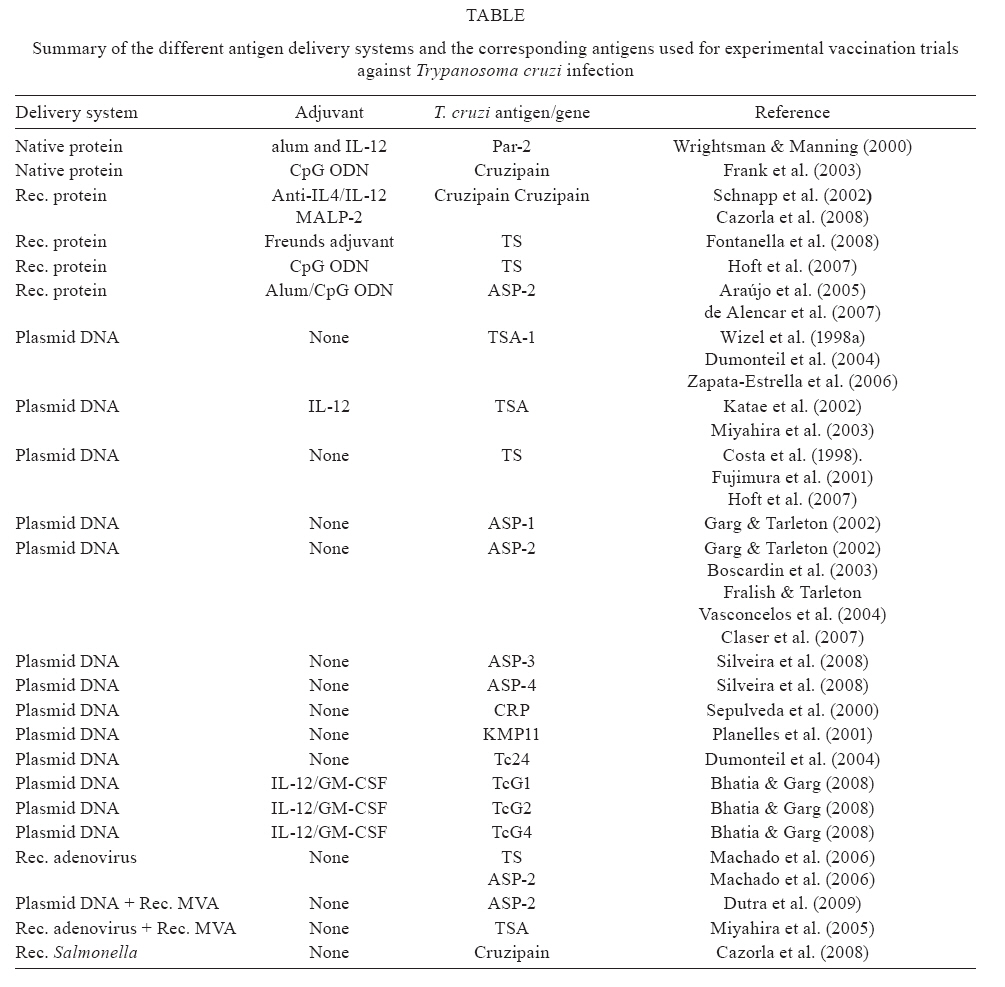
2009 Financial support: FAPESP, INTEV-CNPq (MMR), CNPq (MMR, ELS), FAPESP (BCGA, JRV, CC, FT, FAH, MDR) Code Number: oc09119 ABSTRACT Vaccines have had an unquestionable impact on public health during the last century. The most likely reason for the success of vaccines is the robust protective properties of specific antibodies. However, antibodies exert a strong selective pressure and many microorganisms, such as the obligatory intracellular parasite Trypanosoma cruzi, have been selected to survive in their presence. Although the host develops a strong immune response to T. cruzi, they do not clear the infection and instead progress to the chronic phase of the disease. Parasite persistence during the chronic phase of infection is now considered the main factor contributing to the chronic symptoms of the disease. Based on this finding, containment of parasite growth and survival may be one method to avoid the immunopathology of the chronic phase. In this context, vaccinologists have looked over the past 20 years for other immune effector mechanisms that could eliminate these antibody-resistant pathogens. We and others have tested the hypothesis that non-antibody-mediated cellular immune responses (CD4+ Th1 and CD8+ Tc1 cells) to specific parasite antigens/genes expressed by T. cruzi could indeed be used for the purpose of vaccination. This hypothesis was confirmed in different mouse models, indicating a possible path for vaccine development. Key words: Trypanosoma cruzi - vaccine - immunity The population at risk of infection with obligatory intracellular parasites is estimated to be in the billions in individuals living in tropical and subtropical regions of the world where these parasites are endemic. Although the pattern of disease transmission, infection and mortality caused by these parasites has continued to change over the past 100 years, the global burden of disease is greater than it has ever been. In developing countries, the circumstances supporting disease transmission are unlikely to improve in the next 10 years. In light of these problems, it is essential to develop new strategies for the prevention and control of these diseases. Also, given that global warming will increase, we can anticipate that invertebrate hosts will occupy greater areas of the world and possibly increase transmission of insect-borne diseases. Mass human vaccination against parasitic infection is currently not available. However, the first vaccine against human malaria is scheduled to become commercially available in 2010. Research in this field is considered to be important because in the future, vaccines may complement other strategies such as chemotherapy or vector control for the prevention and control of these parasitic diseases. As a result, immuno-parasitologists have been researching the mechanisms of host-protective immunity and its target antigens for the last 30 years. In spite of the unquestionable impact of vaccination on public health during the last century, only two dozen vaccines are routinely used today. Mechanistically, we do not know how most vaccines provide pre-immunity. The most likely reason for the success of these vaccines is the robust protective properties of specific antibodies. In fact, the efficacy of commercial vaccines has always been measured as a function of the antibody immune response. Because antibodies are indeed so powerful, they exert a strong selective pressure on microorganisms. As expected, many microorganisms have been selected to survive in the presence of antibodies and ultimately use this selection to their advantage. Therefore, in many cases, the "robust" power of specific antibodies is not sufficient to eliminate the pathogen. In this context, over the last 25 years, vaccinologists have looked for other effector mechanisms that could either complement antibodies or act independently to eliminate these antibody-resistant pathogens. The obligatory intracellular parasite Trypanosoma cruzi is responsible for the infection of millions of individuals across the Americas. Although hosts develop a strong immune response to T. cruzi, they do not clear the infection and instead progress to the chronic phase of the disease. Parasite persistence during the chronic phase of infection is now considered to be the main factor contributing to the chronic symptoms of the disease. Based on this finding, containment of parasite growth and survival may be the best route to avoiding chronic phase immunopathology. Prophylactic vaccination would help to reduce or completely eliminate the parasite burden, and thus represents a desirable method to restrict the development of chronic symptoms of the disease. In the past, vaccination studies have failed because researchers have in most cases focused on inducing protective antibodies. Essentially, these studies confirmed that antibodies alone are unable to completely eliminate the parasites and that the antigens expressed in the axenic culture forms of the parasite are not the best targets for protective immunity. These failures have created a paradigm and led researchers to conclude that it would not be possible to generate an anti-T. cruzi vaccine. Over the past 10 years, we and others have challenged this tenet by testing the hypothesis that non-antibody-mediated cellular immune responses to the antigens expressed in the mammalian forms of the parasites could indeed be used for the purpose of vaccination. Using a mouse model of the disease, we confirmed this hypothesis by inducing protective immunity against T. cruzi infection specifically mediated by CD4+ Th1 and CD8+ Tc1 cells specific for antigens expressed by trypomastigotes and amastigotes of T. cruzi. While it is possible to obtain protective immunity against a challenge with a homologous parasite strain, the strain variation observed in certain immunodominant epitopes may impose a severe restriction on the development and use of this method for mass vaccination. This review is not aimed at describing a product that may never exist; instead, our intention is to demonstrate how to elicit immunity to T. cruzi prior to infection in experimental models. CD4+ and CD8+ T cells are important factors in mediating host survival during T. cruzi infection The innate and adaptive immunity induced by T. cruzi is tailored to allow the infection while preventing the death of the host. After contact with the parasite, mammalian hosts initially undergo an acute phase of the infection, characterised by patient parasitemia. This period is critical for both the parasite and the host. The parasite has to concurrently establish a productive infection while at the same time keeping the host alive in order to maintain the parasite's life cycle. After the acute phase, the host's parasitemia levels decrease and the host progresses to the chronic phase of the infection. This latter phase is the period when most transmission to invertebrate hosts occurs. During the early stages of the acute phase, hosts survive because of strong innate immunity, mediated mostly by Toll-like receptors through the activation of the adaptor molecule MyD88 (Campos et al. 2004, Bafica et al. 2006, Bartholomeu et al. 2008). However, to avoid elimination, the parasite manages to significantly delay the onset of the CD8+ T cell-mediated immunity (CMI) response (Martin et al. 2006, Tzelepis et al. 2006, 2007). This manoeuvre is critical for the parasite to establish the infection. Only after the parasitemia reaches a critical point is a powerful CD8+ T CMI response triggered, drastically reducing the parasitemia and securing host survival. This CD8+ T CMI response, and possibly the CD4+ T cells and antibodies, collectively push the mouse into the long chronic phase of infection, during which the parasite is capable of transmission to the invertebrate host. The role of T-cell-mediated adaptive immunity in the naturally acquired response to T. cruzi infection was determined by the use of genetically modified mice that express neither major histocompatibility complex (MHC) class I and II nor CD4 and CD8 molecules. These genetically deficient mouse strains are highly susceptible to acute infection because they are unable to control parasitemia (Rodrigues et al. 2003, Tarleton 2007). While MHC I- and CD8- deficient mice lack only CD8+ T CMI responses, MHC class II or CD4 deficiency leads to a more general failure, with reductions in both the antibody and the CD8 immune responses (Rottenberg et al. 1993, Tarleton 2007, Tzelepis et al. 2007). Therefore, CD4+ T cells may play an extremely important role in generating different mechanisms of protection. However, their function as an effector mechanism during naturally acquired immunity is less clear, as CD8-deficient mice are highly susceptible to infection with most parasite strains (Tarleton 2007, Tzelepis et al. 2007). During human infection, T cells specific for T. cruzi can be detected in most individuals who are in the chronic phase of infection. Although it is difficult to definitively prove that these cells are important to host resistance, it is plausible that they exert an anti-parasitic effect similar to that seen in the mouse model of infection. During the chronic phase of the disease, most individuals have strong CMI to parasite antigens (Morato et al. 1986, Cetron et al. 1993, Dutra et al. 1996, Barros-Mazon et al. 1997, Wizel et al. 1998b, Ribeirão et al. 2000, Fonseca et al. 2005, Albareda et al. 2006, Alvarez et al. 2008). Events causing depression of CMI, such as immuno-suppressive treatment or HIV infection, lead to the relatively fast development of symptomatic forms of the disease in a percentage of chagasic patients (Rocha et al. 1994). In some cases, the reappearance of patient parasitemia characteristic of the acute phase of infection is observed (Sartori et al. 2002). This evidence suggests that CMI may help to maintain the low levels of parasites observed in chronic chagasic patients. Characterization of the in vivo effector mechanisms mediated by protective T cells Although there is evidence showing that T cells participate in the protective immunity against experimental T. cruzi infection, the precise mechanism(s) used by these cells to eliminate T. cruzi has(ve) not been defined. The fact that T cells secrete IFN-γ suggests that this mechanism may play a role in eliminating intracellular forms of the parasite. IFN-γ is an important mediator of naturally-acquired immunity against the infection. Genetically deficient mice that do not express the IFN-γ receptor or Stat4 are unable to control T. cruzi infection (Holscher et al.1998, Tarleton et al. 2000). However, no direct link between IFN-γ secretion by T cells and the in vivo anti-parasitic activity of these cells has been established to date. The IFN-γ-dependent mechanisms are in part mediated by the production of nitric oxide (NO). In the absence of IFN-γ receptors, NO is not produced by macrophages, suggesting that IFN-γ is the main inducer of NO synthase 2 (NOS2) activation. The evidence that NO is critical for eliminating T. cruzi during acute experimental infection was obtained with genetically deficient mice that do not express NOS2. These animals are highly susceptible to infection with different parasite strains (Holscher et al. 1998, Rodrigues et al. 2000). In addition to producing IFN-γ, CD8+ T cells can lyse host cells infected with T. cruzi, or secrete other potentially active mediators such as TNF-αa, granulisin or a number of different chemokines (Munoz-Fernandez et al. 1992, Aliberti et al. 1999, Stenger et al. 1999). In fact, CD8+ T cells specific for amastigote or trypomastigote antigens are capable of lysing non-phagocytic cells infected with T. cruzi in vitro (Wizel et al. 1998b, Low et al. 1998). However, it is unclear whether lysis and DNA degradation of infected target cells by CD8+ T cells can be effective in restricting T. cruzi infection in vivo. Genetically deficient mice that do not make perforin or granzyme B are not more susceptible to infection than wild-type mice after infection with the Brazil strain of T. cruzi (Kumar & Tarleton 1998). However, other T. cruzi strains behave differently and perforin-deficient mice are highly susceptible to infection (Tzelepis et al. 2006). Based on these observations, perforin-dependent and independent mechanism(s) provide resistance against T. cruzi infection in mice. The elucidation of the anti-parasitic mechanisms mediated by T. cruzi-specific CD8+ T cells will require further investigation using more complex experimental models. Target antigens of the protective immune response and design of recombinant subunit vaccines for experimental trials T. cruzi antigens recognised by immune sera from immune or infected humans or animals were the first antigens to be described at the molecular level. This characterisation allowed researchers to conduct studies using recombinant proteins based on isolated, antigen-specific genes. These isolated genes included members of the large trans-sialidase (TS) surface protein family expressed mainly in the infective trypomastigote and amastigote forms of the parasite. The second group of genes belonged to the family of cysteine-proteases (cruzipain) expressed in all of the different forms of the parasite. Other antigens formed a heterogeneous group including molecules such as the flagellar calcium-binding protein, paraflagellar rod protein-2, LYT-1 antigen, Ribosomal Protein L7a-Like Protein and KMP11, among others. Several delivery antigens were used successfully to induce T. cruzi-specific T lymphocytes and protective immunity against an experimental infection. They included recombinant proteins mixed in the presence of distinct adjuvants, plasmid DNA and, more recently, recombinant viruses and bacteria. Table displays a list matching the different antigens with their delivery system. As shown in Table, protective immune response in the mouse model as measured by a reduction in acute phase parasitemia, tissue parasitism and mortality can be elicited by a variety of antigens using distinct delivery systems. In most cases, immunity elicited by these antigens is associated with type I immune response, generated by IFN-γ producing CD4+ and/or CD8+ T cells. In a few cases, some of the mechanisms of induction of protective immunity were investigated. Following intranasal immunisation with (TS) in the presence of the TLR-9 activator CpG ODN, B cells are critical to inducing type I immune response mediated by CD4+ and CD8+ T cells (Hoft et al. 2007). In this same model, the absence of CD4+ or CD8+ T cells also renders the vaccinated animals completely susceptible to infection. Because these animals were genetically deficient, these cells can be required for the induction or the effector phase of the immune response, or both. Similarly, CD8- deficient mice failed to generate protection after immunisation with native Par-2 protein emulsified in CFA or recombinant adenovirus expressing TS or ASP-2 genes (Wrightsman et al. 2002, Machado et al. 2006). The effector mechanisms responsible for the protective immunity have been evaluated in a few experimental models. Upon plasmid immunisation, depletion of either CD4+ or CD8+ T cells completely reversed protective immunity, thus demonstrating a non-overlapping role for these two subpopulations (Katae et al. 2002, Vasconcelos et al. 2004). Following vaccination with recombinant protein of ASP-2 in alum and CpG ODN, only depletion of CD8+, but not CD4+, T cells reversed protective immunity (Araújo et al. 2005). Finally, by using a single T. cruzi epitope recognised by CD8+ T cells, Miyahira et al. (2005) elicited a protective immune response using a heterologous prime-boost strategy with recombinant adenovirus and vaccinia virus. Although only a few experimental systems found type 1 CD4+ and CD8+ T cells to be necessary, these findings confirm the general paradigm that CD4+ and CD8+ T cells do play a key role in CMI. The question as to whether other cells types are also critical for the adaptive immunity induced by these recombinant vaccines requires additional investigation. In addition, it will be important to systematically define the phenotype and function of the specific T cells in order to establish the immunological status of vaccinated individuals prior to their encounter with the pathogen. In spite of clear evidence that immunisation with T. cruzi antigens can provide protective immunity as measured by a reduction in acute phase parasitemia, tissue parasitism and mortality, it is not clear whether immunisation will lead to either remission or a cure of the chronic phase symptoms of the disease. To determine the role of immunisation in reducing chronic phase disease symptoms, a number of experiments using different animal models must be performed. In many of the models described above, tissue inflammation and parasitism in the late chronic phase were significantly reduced following prophylactic vaccination (Garg & Tarleton 2002, Vasconcelos et al. 2004, Araújo et al. 2005). Therefore it is possible that prophylactic vaccinations indeed halt the development of the chronic phase immunopathologies. Nevertheless, the most compelling evidence of a vaccine's ability to reduce the immunopathology was obtained by therapeutic immunisation with T. cruzi genes encoding the TSA and Tc24 genes (Dunonteil et al. 2004). Whether these results are reproducible using different combinations of mouse and parasite strains remains to be seen. Evasion of T-cell immune responses Viruses are the best known class of pathogens that use a variety of mechanisms to evade the immune recognition by MHC class I restricted CD8+ T cells. Some of these strategies may function at the level of antigen presentation by the infected cells such as: (i) downregulation of MHC class I expression, (ii) interference with cytosolic proteolysis, (iii) inhibition of peptide transport to the endoplasmic reticulum (ER), (iv) retention and destruction of MHC class I molecules in the ER and (v) presentation of mutant epitopes that do not bind to MHC I class molecules. Under these circumstances, infected cells have a limited amount of peptide/MHC-class I complexes on their surface, making it difficult for them to be recognised by specific CD8+ T cells. A detailed review and examples of these evasion mechanisms can be found in Antoniou and Powis (2008). Alternatively, escape mechanisms can interfere with the recognition by MHC class I restricted CD8+ T cells. For example, mutant viruses can be selected with amino acid variations that retain binding to MHC class I molecules but result in reduced recognition by T cells or generate antagonistic peptides that inhibit activation of specific T cells by the MHC-peptide complex (Ploegh 1998, Antoniou & Powis 2008). Most recently, the literature has described viral proteins that interfere with the effector mechanisms mediated by T cells. For example, poxviruses encode several soluble cytokine receptors that bind to and block the activity of IFN-α, β and γ, TNF-α, and IL-1. Also, poxviruses express a protein termed crmA that functionally inhibits Asp-specific cysteine proteases (caspases), therefore blocking apoptosis induced by CTL, TNF or Fas (Ploegh 1998). It is not known whether T. cruzi parasites have evolved to employ these immune evasion mechanisms. To date, not a single evasion mechanism has been confirmed in terms of a CD8+ T CMI response to T. cruzi. This lack of information most likely reflects the fact that the CD8+ T CMI response has not been thoroughly investigated. Recently, the identification of CD8+ T cell epitopes derived against proteins of the TS superfamily allowed the comparison of the immunodominant responses resulting from experimental infection in inbred mice following challenge with distinct parasite strains (Martin et al. 2006, Tzelepis et al. 2006, 2007, 2008). CD8+ T cells of a single mouse strain (C57BL/6 mice) infected with different parasite strains recognised distinct peptides as the immunodominant epitope. The diversity of the immunodominant response between strains presents a significant challenge for developing vaccines. A vaccine would require a number of CD8+ epitopes that cover not only the different human MHC haplotypes but also the different parasite strains. Alternatively, the use of sub-dominant epitopes could favour the protective immune responses without requiring so many epitopes (Rodrigues et al. 2009). Finally, the presence of a strong CD8+ T cell immune response may select variants that no longer act as a target for these protective CD8+ T cells as is the case with HCV or HIV (Tester et al. 2005, Watkins 2008). This hypothesis was tested once following vaccination with TS in the presence of CpG ODN. Parasites isolated from immune mice did not show any increase in resistance to anti-TS immune responses, indicating that antigenically distinct forms of the parasites were not selected by the strong protective immune response (Hoft et al. 2007). The question as to whether the absence of selectively imposed variations has an effect on CD8+ T cell immune evasion should be evaluated in more experimental models. Perspectives The knowledge that T cells are essential for efficient immunity to T. cruzi has raised important questions with basic and applied implications. Several gaps in our basic knowledge are expected to be filled in the years to come. Some of the most relevant questions are: (i) how are specific CD8+ T cells induced and maintained in vivo? (ii) What are the main effector mechanisms operating in vivo against the different forms of the parasite? (iii) Are there specific or general mechanisms of immune evasion used by each strain? These questions are even more relevant and much more complex and difficult to study in humans due to the obvious technical limitations. From the applied point of view, it is now clear that a number of different delivery systems can be efficient in terms of the expansion of protective T lymphocytes. Vaccination trials using non-human primates aimed at inducing T CMI have yet to be conducted and should provide important information about the feasibility of this strategy to develop the first vaccines against T. cruzi infection. It is possible that these studies on the immune response elicited following vaccination with recombinant proteins may contribute to the development of new strategies for the prevention or treatment of T. cruzi infection. However, due to the complex nature of this parasitic disease, we believe that it is too early to anticipate the utility of this strategy in humans. REFERENCES
Copyright 2009 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc09119t1.jpg] |
| |||||||||

{kind=link}