 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 104, No. supp 1, July, 2009, pp. 311-318 Ergosterol biosynthesis and drug development for Chagas disease Julio A Urbina Instituto Venezolano
de Investigaciones Científicas, Apartado 21827, 1020 Caracas, Venezuela Received 19 March
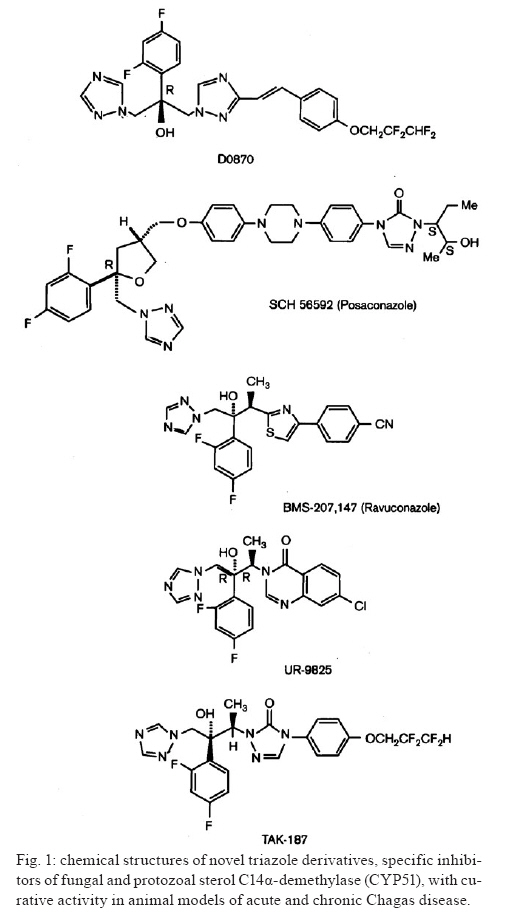
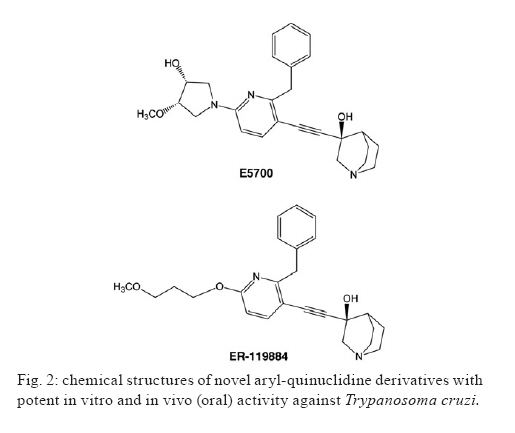
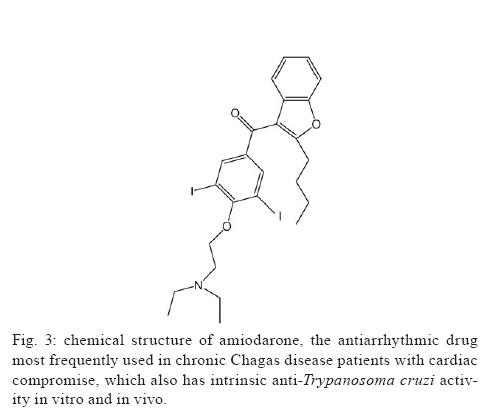
2009 Code Number: oc09123 ABSTRACT This article presents an overview of the currently available drugs nifurtimox (NFX) and benznidazole (BZN) used against Trypanosoma cruzi, the aetiological agent of Chagas disease; herein we discuss their limitations along with potential alternatives with a focus on ergosterol biosynthesis inhibitors (EBI). These compounds are currently the most advanced candidates for new anti-T. cruzi agents given that they block de novo production of 24-alkyl-sterols, which are essential for parasite survival and cannot be replaced by a host's own cholesterol. Among these compounds, new triazole derivatives that inhibit the parasite's C14α sterol demethylase are the most promising, as they have been shown to have curative activity in murine models of acute and chronic Chagas disease and are active against NFX and BZN-resistant T. cruzi strains; among this class of compounds, posaconazole (Schering-Plough Research Institute) and ravuconazole (Eisai Company) are poised for clinical trials in Chagas disease patients in the short term. Other T. cruzi-specific EBI, with in vitro and in vivo potency, include squalene synthase, lanosterol synthase and squalene epoxidase-inhibitors as well as compounds with dual mechanisms of action (ergosterol biosynthesis inhibition and free radical generation), but they are less advanced in their development process. The main putative advantages of EBI over currently available therapies include their higher potency and selectivity in both acute and chronic infections, activity against NFX and BZN-resistant T. cruzi strains, and much better tolerability and safety profiles. Limitations may include complexity and cost of manufacture of the new compounds. As for any new drug, such compounds will require extensive clinical testing before being introduced for clinical use, and the complexity of such studies, particularly in chronic patients, will be compounded by the current limitations in the verification of true parasitological cures for T. cruzi infections. Key words: chemotherapy - Trypanosoma cruzi - sterol biosynthesis - sterol C14α demethylase - squalene synthase - oxidosqualene cyclise - amiodarone Although a century has passed since the discovery of American Trypanosomiasis or Chagas disease (Chagas 1911), it remains the largest parasitic disease burden on the American continent, yet it is one of the most neglected parasitic diseases in the world; to combat the disease requires an integrated approach that addresses its underlying eco-socio-economic causes (Morel et al. 2005, Dias 2007, Holveck et al. 2007, Hotez et al. 2008). Significant advances have taken place to control the vectorial and transfusional transmission of the disease in some parts of the continent. In particular, the Southern Cone initiative has successfully interrupted reservoir-to-human and human-to-human propagation of the disease in Uruguay, Chile and Brazil in recent years and has led to a marked reduction of the overall prevalence and the population at risk (Dias et al. 2002, Schofield et al. 2006). However, many challenges remain before full control or elimination of the disease can be achieved among them, inequality of the control programs in the various endemic areas (Attaran 2006, Schofield et al. 2006, Aguilar et al. 2007, Guhl 2007), limitations of currently available specific chemotherapy, and insufficient treatment coverage for those currently infected with the aetiological agent, the parasitic protozoan Trypanosoma cruzi (Urbina & Docampo 2003, Pinto Dias 2006, Jannin & Villa 2007, Tarleton et al. 2007, Villa et al. 2007, Rassi et al. 2009). Relevance of specific chemotherapy for Chagas disease and limitations of currently available drugs Although the role of T. cruzi in the pathology and treatment of the acute phase of Chagas disease is widely accepted (Brener & Gazzinelli 1997, Bahia-Oliveira et al. 2000, Kirchhoff et al. 2004), the parasite's influence on the pathogenesis of chronic Chagas disease has been the subject of controversies for decades (Cunha-Neto et al. 1995, Kalil & Cunha-Neto 1996, Tarleton & Zhang 1999, Tarleton 2001, 2003). A key component of Chagas disease pathology is a sustained and diffuse inflammation of the affected organs with associated cytolysis and fibrosis, and the recognition of this phenomenon has led to intense studies to elucidate the origin and maintenance of such inflammatory responses. Several studies have implicated autoimmunity as a primary factor leading to the persistent inflammation associated with chronic Chagas disease pathology including chronic Chagas' cardiomyopathy (Cunha-Neto et al. 1995, Kalil & Cunha-Neto 1996). This hypothesis was based on the apparent absence of the parasite in the characteristic inflammatory lesions of the heart and gastrointestinal tract coupled with the presence of "anti-self" antibody responses in Chagas disease patients; the latter is postulated to result from molecular mimicry between parasite antigens and a host's cellular components (Cunha-Neto et al. 1995, Kalil & Cunha-Neto 1996). According to such a hypothesis, after the parasite triggers the autoimmune response in the host, its persistence does not play a pivotal role in the pathogenesis of the disease, and even successful antiparasitic treatment may not lead to an improvement in the clinical outcome of the patients. In fact, the autoimmune hypothesis of chronic Chagas disease pathogenesis, which temporally followed Köberle's neurogenic postulate, stalled for decades the development of new specific chemotherapeutic approaches for this disease, as antiparasitic treatment was considered irrelevant in both aforementioned hypotheses (Urbina 1999b, Urbina & Docampo 2003). However, more recent studies using more sensitive methodologies have shown clear correlations between the presence of the parasite and the inflammatory processes that underlie the pathological processes associated with chronic Chagas disease (Tarleton et al. 1997, Tarleton & Zhang 1999, Tarleton 2001, 2003, Marin-Neto et al. 2007). This fact, coupled with the finding that T. cruzi-specific CD8+ and CD4+ T cells, are consistently associated with inflammatory infiltrates rich in Th1 cytokines, has led to the notion that parasite persistence is a necessary and sufficient condition to generate and sustain a Th1-biased inflammatory response in infected tissues, which may include autoimmune phenomena (Tarleton et al. 1997, Tarleton & Zhang 1999, Tarleton 2001, 2003, Marin-Neto et al. 2007). Thus, there is a growing consensus that the persistence of parasites, along with unbalanced immune responses in some individuals, leads to a sustained inflammatory response that underlies the characteristic lesions of chronic Chagas disease. This new paradigm suggests, in contrast to long-held views, that eradication of T. cruzi may be a prerequisite to arrest the evolution of Chagas disease and avert its irreversible long-term consequences, and it implies that this condition must be treated primarily as an infectious, not autoimmune, condition (Tarleton 2001, Urbina & Docampo 2003, Marin-Neto et al. 2007, Tarleton et al. 2007, Rassi et al. 2009). Current specific chemotherapy for Chagas disease relies on two compounds empirically introduced for clinical use in the late 1960's and early 1970's: nifurtimox (NFX) [Lampit®, Bayer; 5-nitrofuran 3-methyl-4-(5'-nitrofurfurylideneamine) tetrahydro-4H-1,4-tiazine-1,1-dioxide] and benznidazole (BZN) (Rochagan®, Radanil®, Roche; N-benzyl-2-nitroimidazole acetamide) (Coura & de Castro 2002, Pinto Dias 2006). Numerous clinical studies, admittedly of unequal quality and duration, have shown that both drugs have significant activity during the acute phase (up to 80% negativisation of all parasitological and conventional serological tests) (Cançado 1999, 2002). However, their efficacies vary according to geographical area, most likely due to differences in drug susceptibility among different T. cruzi strains (Filardi & Brener 1987, Murta et al. 1998, Kirchhoff 1999). Moreover, it has been reported in the last 15 years that BZN also exhibits significant curative activity in those with recent onset (up to a few years) of chronic disease; 60-70% of radical parasitological cures have been observed among children up to 14 years of age in Brazil and Argentina (de Andrade et al. 1996, Andrade & Zicker 1997, Sosa Estani et al. 1998, Sosa Estani & Segura 1999, Andrade et al. 2004). On the other hand, other studies in patients of the same age range treated with either BZN or NFX contradict these conclusions (Silveira et al. 2000, Solari et al. 2001). The major limitation of these compounds is insufficient antiparasitic activity in the chronic form of the disease, i.e., > 80% of treated patients are not parasitologically cured according to the classical criteria indicated above for acute infections (Cançado 2002). These results have now been confirmed using PCR-based methods in both humans and experimental animals (Anez et al. 1999, Braga et al. 2000, Lauria-Pires et al. 2000, Britto et al. 2001, Garcia et al. 2005, Martins et al. 2008, Fernandes et al. 2009). The reasons for the marked difference in the antiparasitic efficacy of nitro-heterocyclic compounds in the acute and chronic stages of the disease are unclear (Cançado 2002), but they may be related to unfavourable pharmacokinetic properties, such as relatively short half-lives and limited tissue penetration (Raaflaub & Ziegler 1979, Raaflaub 1980, Workman et al. 1984), which will limit their action in the chronic stage when the parasites are mostly confined in deep tissues and undergo slow replication (Urbina 1999a, Urbina & Docampo 2003). In any case, verifying a true parasitological cure in chronic infection, where the levels of circulating parasites can be extremely low or undetectable even with the most sensitive PCR methods available (Martins et al. 2008), remains an extremely challenging problem that requires developing other criteria, such as immunological assays that measure T-cell responses to drug treatment (Bustamante et al. 2008). Nevertheless, several observational clinical studies have shown that chronic patients subjected to antiparasitic treatment with BZN, although not parasitologically cured, achieve a significant reduction in the occurrence of electrocardiographic changes and a lower frequency of deterioration of their clinical condition (Viotti et al. 1994, 2006, Fragata Filho et al. 1995, Bahia-Oliveira et al. 2000, Fabbro De Suasnabar et al. 2000, Sosa-Estani & Segura 2006, Viotti & Vigliano 2007); however, other studies do not confirm these findings (Lauria-Pires et al. 2000, Sosa-Estani & Segura 2006). The positive effect of current specific treatments, despite their inability to eradicate the parasite, on clinical evolution has been explained in terms of the parasite persistence hypothesis. That is, a drug-induced reduction of the parasite load in infected tissues reduces the severity of the associated inflammatory processes (Tarleton 2001, Viotti et al. 2006, Tarleton et al. 2007, Viotti & Vigliano 2007). Unfortunately, both drugs have unwanted side effects that can lead to treatment discontinuation (Coura & de Castro 2002, Rassi et al. 2009). The incidence of such side effects is variable and apparently depends on age, geographic region and the quality of the clinical supervision of the treatment (Coura & de Castro 2002). Moreover, both NFX and BZN are genotoxic, as expected form their chemical nature and reactivity. Due to the empirical nature of the discovery and development of NFX and BZN, these drugs were registered and used in clinical settings for many years without a clear understanding of their mechanism of action. By the mid-1980's, basic studies began to illuminate the molecular basis of both their anti-T. cruzi activity and toxicity (Docampo & Moreno 1986, Docampo 1990). It has been experimentally demonstrated that the antiparasitic action of NFX is associated with the production of large amounts of toxic reactive oxygen species (ROS, such as superoxide anion and hydrogen peroxide), resulting from the reaction of oxygen with unstable nitroanion metabolites (Docampo & Moreno 1986). However, BZN cytotoxicity is not associated with the generation of significant levels of ROS; instead, it probably results from reductive stress, which involves covalent modification of macromolecules by nitroreduction intermediates (Docampo & Moreno 1986, Docampo 1990). Given the significant limitations of the currently available drugs, particularly for the treatment of chronic disease, new approaches to specific chemotherapy of Chagas disease have been advanced in the last two decades, and these will be discussed in the following sections with a focus on those candidates with the greatest probability of entering clinical studies in the short to medium-term. Ergosterol biosynthesis inhibitors (EBI) as potential new therapeutic agents for Chagas disease C14α-lanosterol demethylase (CYP51) inhibitors - Research during the last two decades has consistently demonstrated that T. cruzi, like most fungi and yeasts, requires specific sterols for cell viability and proliferation in all stages of its life cycle. In particular, the ergosterol biosynthesis pathway has been chemically validated in vitro at many different steps (Urbina 2002, Urbina & Docampo 2003). Yet, several studies have shown that commercially available EBI, which are highly successful for the treatment of fungal diseases (such as ketoconazole, itraconazole or terbinafine), have suppressive but not curative effects against T. cruzi infections in humans or experimental animals and are unable stop the progression of the disease (Urbina 2002, Urbina & Docampo 2003). However, over the past 15 years, new triazole derivatives, e.g., D0870 (Zeneca Pharmaceuticals) and posaconazole (POS; SCH 56592; Schering-Plough Research Institute, SPRI) (Fig. 1), which are potent and selective inhibitors of fungal and protozoan cytochrome P-450-dependent CYP51, have been found to induce radical parasitological cure in murine models of acute and chronic Chagas disease (Urbina et al. 1996, Urbina 2002, Urbina & Docampo 2003). These were the first compounds reported to display curative activity in both forms of the disease. Furthermore, such compounds were able to eradicate nitrofuran and nitroimidazole-resistant T. cruzi strains from infected mice, even if the hosts were immunosuppressed (Urbina 2002, Urbina & Docampo 2003). It has been argued that the remarkable in vivo antiparasitic activities of these triazole derivatives result from a combination of their potent and selective intrinsic anti-T. cruzi activity (the minimal growth inhibitory concentrations against the intracellular amastigote form is in the nanomolar to sub-nanomolar range) and special pharmacokinetic properties (long terminal half-life and large volumes of distribution) (Urbina 1999a, 2002, Urbina & Docampo 2003). More recent studies with POS, a structural analogue of itraconazole, have shown that this compound can eradicate the intracellular amastigote forms from cultured cardiomyocytes while allowing the full reassembly of the host cell's cytoskeleton and contractile apparatus (Silva et al. 2006). Other studies have demonstrated that the anti-T. cruzi activity of POS in a murine model of acute Chagas disease is much less dependent on interferon-γ than that of BZN (Ferraz et al. 2007). It has also been shown that ablation of TCD4+, TCD8+ and B lymphocytes has distinct effects on POS and BZN activity in the same experimental model (Ferraz et al. 2009). POS efficacy was especially dependent on the presence of functional TCD8+ cells, but relatively insensitive to the absence of LB cells, yet the reverse was true for BZN; the activity of both drugs was markedly reduced in the absence of TCD4+ cells. These results were interpreted in terms of the different parasite stages preferentially targeted by the two drugs (intracellular amastigotes by POS, extracellular trypomastigotes by BZN) and distinct cooperation patterns with the host's immune system (Ferraz et al. 2009). An independent study in a similar experimental model found that POS was more effective than BZN in preventing heart damage and promoting a trypanocidal immune response (Olivieri et al., unpublished observations). POS was registered in 2005 in the European Union and Australia for treatment and, in 2006, in the USA for the prophylaxis of invasive fungal infections and for the treatment azole-resistant candidiasis. It is poised to enter clinical development for the treatment of human chronic Chagas disease in the next 12 months. This will be the first rationally designed drug to enter clinical trials for the treatment of this disease. Other triazoles (Fig. 1), such as TAK-187 (Takeda Chemical Company) (Urbina et al. 2003c, Corrales et al. 2005), UR-9825 (Uriach & Company) (Urbina et al. 2000, Guedes et al. 2004) and ravuconazole (ER-30346, Eisai Co.; BMS 207,147; Bristol-Myers Squibb) (Urbina et al. 2003b) also exhibit trypanocidal activity in vitro and in vivo. TAK-187 is a long-lasting triazole derivative with broad-spectrum antifungal activity, which also possesses very potent anti-T. cruzi activity in vitro and is capable of curing both acute and chronic infections in murine hosts even when the infecting strain is resistant to nitrofuran and nitroimidazole (Urbina et al. 2003c). More recent work has shown that this compound is superior to BZN in preventing cardiac damage in a murine Chagas disease model (Corrales et al. 2005). UR-9825 is another potent fungal and protozoan CYP51 inhibitor with remarkable in vitro anti-T. cruzi activity (Urbina et al. 2000). However, its very short half-life in the mouse (< 0.5 h) precluded in vivo studies in this animal model, but work in a canine model has demonstrated that the compound exhibits curative activity against established infections of the virulent Y strain of T. cruzi with very low toxicity, although drug resistance was encountered with the Berenice-78 strain (Guedes et al. 2004). Finally, ravuconazole has also been shown to be very active against T. cruzi in vitro, but its in vivo activity in mice was limited, most likely due to inadequate pharmacokinetic properties in this animal model (terminal half-life of 4.5 h) (Urbina et al. 2003b). Similarly, the activity of ravuconazole in a canine model of acute Chagas disease was found to be suppressive, not curative, a result that was again attributed to the relatively short half-life of the compound in dogs (8.8 h) (MT Bahia & JA Urbina, unpublished observations). However, these results do not necessarily rule out the potential usefulness of this compound in the treatment of human T. cruzi infections since its minimal inhibitory concentration against intracellular amastigotes (1 nM) is 1,000-5,000-fold lower than the levels attainable in human plasma following multiple oral dosages, and its terminal half-life in man is > 120 h (Mikamo et al. 2002, Andes et al. 2003). This compound is another prime candidate for clinical trials regarding Chagas disease patients. Another class of azole-based CYP51 inhibitors that possess potent anti-T. cruzi activity in vitro and in vivo was serendipitously discovered in the course of a research program to identify parasite-specific protein farnesylation inhibitors and represents an interesting alternative to the proprietary compounds described above (Buckner et al. 2003, Hucke et al. 2005, Buckner 2008, Kraus et al. 2009). Very recently, a new class of T. cruzi CYP51 inhibitors, based on the N-[4-pyridyl]-formamide moiety and derived from a Mycobacterium tuberculosis screen hit, has been described (Chen et al. 2009); one of the new compounds was able to eradicate intracellular amastigotes grown in cultured mouse macrophages. Squalene synthase (SQS) inhibitors - A different group of promising T. cruzi agents among EBIs is that of SQS inhibitors. SQS catalyses the first committed step in sterol biosynthesis and has been the subject of intense study by both academic and industrial groups as an attractive target for cholesterol lowering agents. It presents potentially significant advantages over currently available statins (Tansey & Shechter 2001, Menys & Durrington 2003, Suckling 2006, Charlton-Menys & Durrington 2007). This enzyme has been recently chemically validated as a chemotherapeutic target in T. cruzi and Leishmania mexicana (Urbina et al. 2002). Further studies led to the discovery that E5700 and ER-119884, two novel quinuclidine SQS inhibitors currently under development as cholesterol and triglyceride lowering agents in humans by Eisai Company (Fig. 2), have very potent anti-T. cruzi activity in vitro. One of them (E5700) was able to provide full protection against death, and it completely arrested development of parasitaemia in a murine model of acute disease when given orally (Urbina et al. 2004), this was the first report of an orally-active SQS inhibitor as an anti-infective agent. Although these compounds and other aryl-quinuclidines are also potent inhibitors of mammalian SQS (McTaggart et al. 1996, Ward et al. 1996, Ishihara et al. 2003, 2004), their selective antiparasitic activity in vitro and in vivo might be explained by the capacity of a host's cells to compensate for the blockade of de novo cholesterol synthesis by up-regulating the expression of LDL receptors and capturing this sterol from the growth medium or serum (Goldstein & Brown 2001). In contrast, there is no way for the parasite to compensate in this manner for the quinuclidine-induced blockade of ergosterol biosynthesis since there are no appreciable amounts of ergosterol in host cells or growth media (Urbina et al. 2002, 2004). However, the requirement of some key organs (such as testis) of an elevated "endogenous" cholesterol supply could pose a significant limitation for the prolonged use of these SQS inhibitors. In fact, prolonged use of selective SQS inhibitors (and statins) has been associated with testicular atrophy in experimental animals (Eisai Co, data on file). Thus, the use of this class of compounds as antiparasitic agents will probably require parasite-specific SQS inhibitors in much the same way that current antifungal azoles are specific for fungal and parasite CYP51 while having much less of an effect on the enzyme's mammalian orthologue. Recent work has demonstrated progress towards this goal, as the gene encoding T. cruzi SQS has been cloned and expressed in Escheria coli, resulting in the production of a soluble, fully active, recombinant enzyme, which has been used to identify parasite-specific SQS inhibitors (Orenes Lorente et al. 2005, Sealey-Cardona et al. 2007). Finally, it has also been shown that aryloxylethyl thiocyanates such as WC-9 (4-phenoxyphenoxyethylthiocyanate), a new class of potent and selective anti-T. cruzi agents (Szajnman et al. 2000, Elhalem et al. 2002), act by selectively inhibiting the parasite's SQS (Urbina et al. 2003a). Lanosterol synthase [oxidosqualene cyclise (OSC)] inhibitors - Another advance in the area of EBIs as antiparasitic agents is the validation of OSC as a chemotherapeutic target in T. cruzi and related parasites (Buckner et al. 2000, 2001, Joubert et al. 2001). Buckner et al. demonstrated the potent and selective in vitro antiparasitic activities of OSC inhibitors (Buckner et al. 2001, Hinshaw et al. 2003) and a patent submitted by the same authors claims the use of OSC inhibitors as chemotherapeutic agents for the specific treatment of parasitic infections, including Chagas disease [US Patent WO0076316 (Urbina 2003)]. However, no in vivo activity of this class of compounds has been reported. A related development was the surprising discovery that amiodarone, a bis-aryl-ketone (Fig. 3) used extensively as an antiarrhythmic drug in chronic Chagas disease patients with cardiac compromise (Rosenbaum et al. 1983, Rassi et al. 2000, 2009), has intrinsic anti-T. cruzi activity in vitro and in vivo and that combinations of this drug with posaconazole have synergistic effects (Benaim et al. 2006). It was found that amiodarone functions via dual mechanisms of action against this parasite: disruption of Ca2+ homeostasis and blockade of de novo ergosterol biosynthesis at the level of OSC, which explains the synergistic effects observed with posaconazole. The results indicate that Chagas disease patients being treated with amiodarone may have the added benefit of a reduction of their parasite burden and enhancement of the effects of antiparasitic treatment (Benaim et al. 2006). Hybrid compounds with dual mechanisms of action - An interesting development in this area is a recent report on the synthesis and chemical/biological characterisation of compounds with dual mechanisms of action resulting from the presence of two pharmacophores in the same molecule: a nitrofuran moiety that leads to free radical generation and redox cycling and a heteroallyl moiety that inhibits squalene epoxidase, a key enzyme in sterol biosynthesis (Gerpe et al. 2008). These heteroallyl-containing 5-nitrofuran compounds were very active against both the extracellular and intracellular forms of the parasite in vitro and more potent than NFX and terbinafine, a commercial squalene epoxidase inhibitor with antifungal activity. The potential of this new class of compounds is underscored by previous studies (Araujo et al. 2000) that showed that combination therapy with BZN and the EBI ketoconazole had synergistic effects in a murine model of acute Chagas disease. In conclusion, EBI are currently the most advanced candidates for new specific treatments for Chagas disease, as some of them could enter clinical trials in the short-term (within 24 months). Their primary advantages over currently available therapies include their higher potency in both acute and chronic infections, activity against NFX and BZN-resistant T. cruzi strains, and their enhanced tolerability and safety profiles resulting from their selective mechanisms of action. Limitations may include complexity and cost of manufacturing the new compounds and the uneven advancement of various candidates in the drug development process, but there are several options in the pipeline that can address such issues. As for any new drug, such compounds will require extensive clinical testing before being introduced for clinical use. The complexity of these studies, particularly in chronic patients, will be compounded by the current limitations in the verification of true parasitological cures for T. cruzi infections. Despite these shortcomings, this class of compounds remains the best hope for millions of patients currently living with this insidious, silent killer. REFERENCES
Copyright 2009 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc09123f1.jpg] [oc09123f3.jpg] [oc09123f2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}