 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 104, No. 4, July, 2009, pp. 583-591 ARTICLES Circulating natural killer and γδ T cells decrease soon after infection of rhesus macaques with lymphocytic choriomeningitis virus Juan D RodasI; Cristiana CairoII; Mahmoud DjavaniII; Juan Carlos ZapataII; Tracy RuckwardtIII; Joseph BryantII; C David PauzaII; Igor S LukashevichII; Maria S SalvatoII, + IGrupo
de Investigaciones en Ciencias Veterinarias Centauro, Facultad de Ciencias Agrarias,
Universidad de Antioquia, Medellín, Colombia Financial support: National Institutes of Health (AI5252367 to ISL, RR138980 to ISL, AI53620 to MSS, AI53619 to MSS) Received 4 November
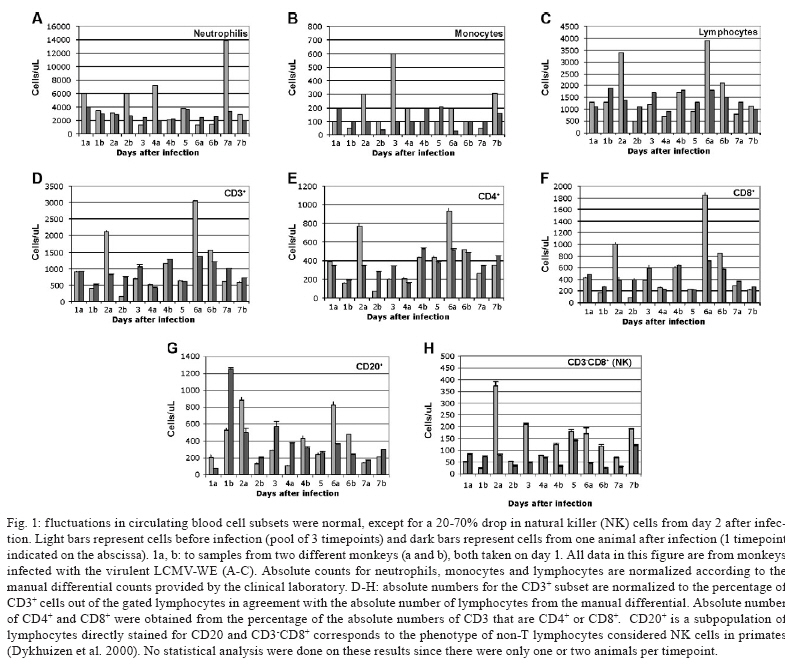
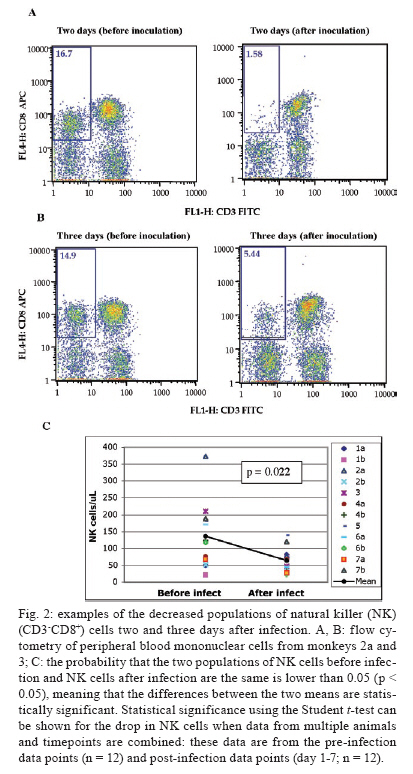
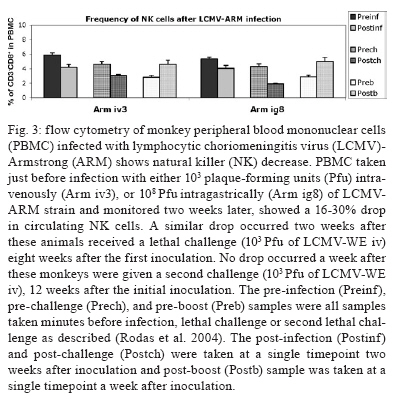
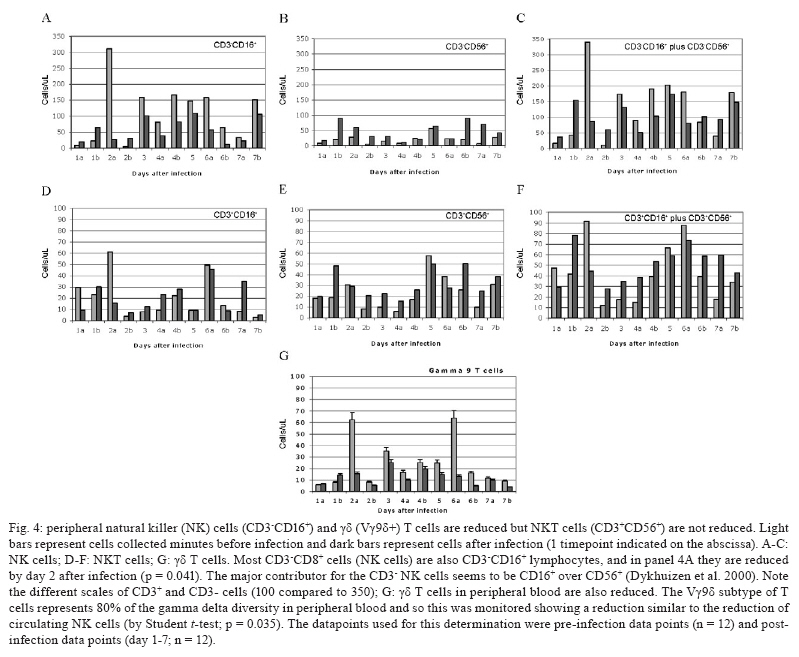
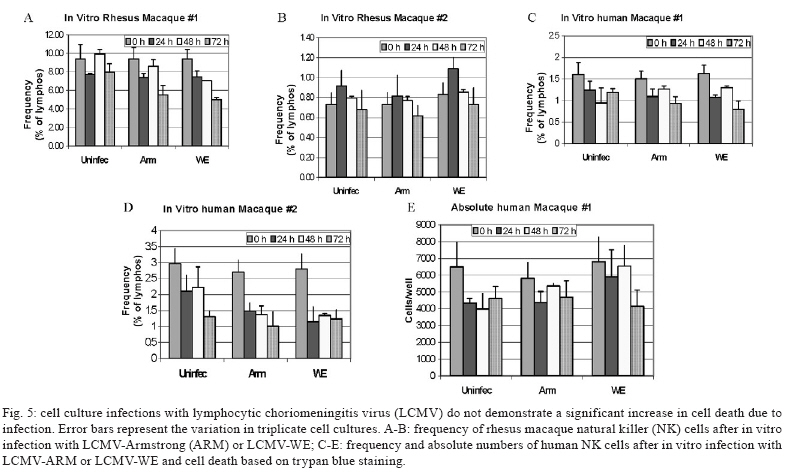
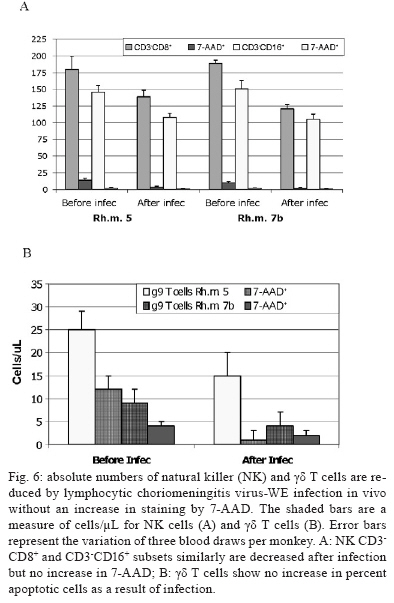
2008 Code Number: oc09134 ABSTRACT Rhesus macaques infected with the WE strain of lymphocytic choriomeningitis virus (LCMV-WE) serve as a model for human infection with Lassa fever virus. To identify the earliest events of acute infection, rhesus macaques were monitored immediately after lethal infection for changes in peripheral blood mononuclear cells (PBMCs). Changes in CD3, CD4, CD8 and CD20 subsets did not vary outside the normal fluctuations of these blood cell populations; however, natural killer (NK) and γδ T cells increased slightly on day 1 and then decreased significantly after two days. The NK subsets responsible for the decrease were primarily CD3-CD8+ or CD3-CD16+ and not the NKT (primarily CD3+CD56+) subset. Macaques infected with a non-virulent arenavirus, LCMV-Armstrong, showed a similar drop in circulating NK and γδ T cells, indicating that this is not a pathogenic event. Vγ9 T cells, representing the majority of circulating γδ T cells in rhesus macaques, displayed significant apoptosis when incubated with LCMV in cell culture; however, the low amount of cell death for virus-co-cultured NK cells was insufficient to account for the observed disappearance of this subset. Our observations in primates are similar to those seen in LCMV-infected mice, where decreased circulating NK cells were attributed to margination and cell death. Thus, the disappearance of these cells during acute hemorrhagic fever in rhesus macaques may be a cytokine-induced lymphopenia common to many virus infections. Keywords: NK cells - γδ T - rhesus macaque - LCMV - hemorrhagic fever Arenaviruses are rodent-borne pathogens that have occasionally been known to cause lethal diseases in human beings (Oldstone 2002, Salvato & Rodas 2005). Every year, Lassa fever and the South American hemorrhagic fever viruses account for almost half a million cases worldwide, with approximately 16% mortality (Jahrling et al. 1985, McCormick et al. 1986, 1987, Fisher-Hoch et al. 2000). Although rodents serve as a reservoir for arenaviruses, studies of hemorrhagic fever have relied on guinea pig, hamster and primate models rather than mouse models because the disease mechanism is fundamentally different in non-reservoir species (Peters et al. 1987). Numerous murine studies of viral persistence and cell-mediated immunity have involved the prototype arenavirus, lymphocytic choriomeningitis virus (LCMV) (Zinkernagel & Doherty 1974, Ahmed et al. 1984, Salvato et al. 1991). The WE strain of LCMV is hepatotropic in mice, guinea pigs and primates (Riviere et al. 1985, Zinkernagel et al. 1986, Lukashevich etal. 2002, 2003), but in contrast to murine LCMV infections, LCMV-WE infection of rhesus macaques can resemble Lassa hemorrhagic fever in human beings (Jahrling et al. 1980, Peters et al. 1987, Lukashevich et al. 2002, 2003). The Armstrong (ARM) strain of LCMV does not cause overt disease in monkeys, even after intravenous inoculation (Danes et al. 1963, Peters et al. 1987, Lukashevich et al. 2002, 2004). Several immunological studies have shown the importance of CD8 T lymphocytes in both protection and in induction of immuno-pathogenesis in the mouse model (Doherty & Zinkernagel 1974, Zinkernagel & Doherty 1974, Bonilla et al. 2002). More recently, we demonstrated the importance of cell-mediated immunity in survival after a lethal intravenous challenge in the primate model (Rodas et al. 2004). However, no studies in primates have yet described the earliest changes in peripheral blood subsets after LCMV infection. Significant changes were not observed in most PBMC subsets, but we noted changes in circulating natural killer (NK) and γδ T cells. Although these subsets represent less than 5% (NK) and less than 1% (NKT and γδ T cells) of circulating PBMCs, they are powerful effectors of innate immunity and the subsequent acquired immune responses to viral infection (Poccia et al. 2002, Jacobs et al. 2005). Although our goal was initially to identify early changes that discriminate the Lassa-like disease from a benign infection, it appears that the NK-cell depletion occurs after infection with both virulent and non-virulent viruses and could be a manifestation of the cytokine-mediated lymphopenia previously described for several viruses (McNally et al. 2001, Peacock et al. 2003). This study describes a 20-70% decrease in circulating NK and γδ cells and explores some possible explanations for these observations. MATERIALS AND METHODS Virus stocks and cell culture - Stocks of serum-free LCMV were produced in Vero E6 cells and stored at 107 to 108 plaque-forming units (pfu)/mL for use in all monkey inoculations. The LCMV-WE and LCMV-ARM strains have been well characterized (Salvato & Shimomaye 1989, Djavani et al. 1998) and their use in monkeys has been previously described by our group. A dose of 103 pfu LCMV-WE inoculated intravenously in unvaccinated macaques is uniformly lethal (Lukashevich et al. 2002, 2003, 2004, Rodas et al. 2004). Rhesus macaque inoculations and challenges - Rhesus monkeys were housed in a BSL-2/3 facility. The 12 animals used in this study were all 2-5-year-old females with normal weights and activity levels. All monkeys were first anesthetized (ketamine, 20 mg/kg) and then intravenously inoculated (via the left saphenous vein) with a lethal dose (103 pfu) of LCMV-WE in 0.5 mL of PBS. Animals were observed and sacrificed at the rate of one or two per day during the first week after infection. Two additional animals, Rh-iv3 (ARM) and Rh-ig8 (ARM), were infected with LCMV-ARM as previously described (Rodas et al. 2004). All experimental procedures and protocols were reviewed and approved by the Institutional Animal Care and Use Committee. The methods were consistent with the recommendations of Panel on Euthanasia of the American Veterinary Medical Association. Necropsy was conducted in a negative-pressure, Hepa-filtered room and protective gear included respirators (Hepa 12 Air-supply units; Lab Safety Supply #0E-67642, Janesville, WI) and barrier clothing (i.e., disposable Tyvek jump suits, boots, gloves and face shields; Fisher Scientific, Pittsburgh, PA). Blood collection - Two blood samples were taken per animal, one before infection and one just before euthanasia. Samples were submitted to the clinical laboratory for complete blood counts. Peripheral blood mononuclear cells (PBMCs) were obtained from anticoagulant-treated blood and centrifuged over Ficoll-Paque (Amersham Biosciences) as described elsewhere (Pauza et al. 1997). Other human and monkey samples were obtained from healthy donors at the Institute of Human Virology. Analysis of lymphocyte subsets and apoptosis of infected rhesus macaques - Flow cytometry was used to monitor changes in white blood cell subsets after infection. PBMCs were paraformaldehyde-fixed and stained with anti-human monoclonal antibodies known to cross-react with rhesus CD3+, CD4+, CD8+, CD16+, CD20+, CD56+ and Vγ9 T-cell receptor (TCR) lymphocyte subsets. NK cells are considered CD3-CD8+ because 99% of these cells are CD16+ or CD56+ (classical NK markers) (Dykhuizen et al. 2000). FITC-conjugated monoclonal antibodies against CD3, CD20 (Becton-Dickinson, Mountain View, CA) and Vγ9 TCR (Beckman Coulter, Miami, FL), a PerCP-conjugated monoclonal antibody against CD4, an APC-conjugated monoclonal antibody against CD8 and PE-conjugated monoclonal antibodies against CD16 and CD56 (Becton-Dickinson) were used with relevant isotype controls as described previously (Pauza et al. 1997, Lukashevich et al. 2002). Samples were analyzed on a FACScan flow cytometer (Becton-Dickinson) and data were processed using FlowJo 2.7.8 software (Three star, 1997-1998). Absolute counts were determined by multiplying the percentage for each subset by the absolute lymphocyte count obtained from clinical hematology data. Apoptosis induced by viral infection was detected by staining with 7-amino actinomycin D (7-AAD) as described previously (Lecoeur et al. 1997). Briefly, 2.5 x 105 PBMCs were labelled for surface markers and then incubated for 20 min at 4°C in PBS that contained 20 μg/mL of 7-AAD (Sigma, St. Louis, MO). Samples were washed in PBS + 2% FBS containing 20 μg/mL of non-fluorescent actinomycin D (AD, Sigma) and fixed in the same buffer containing 1% paraformaldehyde. Samples were analyzed 15 min later (10,000 events/sample) using the FL-3 channel to detect 7-AAD staining. Analysis of apoptosis and lymphocyte subsets after culturing PBMCs with or without virus - Blood samples were taken from healthy monkey and human donors and cultured in the presence or absence of LCMV and/or IL-2, to determine the level of cell survival after 24, 48 or 72 h. Cells were monitored by trypan blue and 7-AAD staining. The cells from the human samples were counted at every time point to determine the absolute counts for each cell subset. In all of the cases where error bars are depicted, samples were run in triplicate to estimate the average and standard deviation for each time point and treatment. Detection of infectious virus in plasma - LCMV-WE was detected in plasma by plaque assay as described (Lukashevich et al. 2002). In brief, 10-fold dilutions of plasma samples were added to monolayers of Vero cells (about 2.5 x 105 cells/well) in 6-well plates and incubated for 1 h at 37°C. After incubation, wells were overlaid with 1% agarose in MEM + 2% FBS and incubated again at 37°C. After five days, the wells were treated with formaldehyde for 30 min. The agarose overlay was then removed and monolayers were stained with crystal violet, washed with tap water and dried and visible plaques were counted to estimate the pfu/mL of virus in plasma. RT-PCR for detection of viral RNA - RNA was extracted from PBMCs or tissue samples using the Trizol reagent (Invitrogen, Carlsbad, CA). Isolated RNA was treated with DNAse (Promega, Madison, WI) 1 unit/μg, at 37°C for 30 min and purified again with the RNAeasy Mini Kit (Qiagen, Valencia, CA). RNA was converted to cDNA with the avian myeloblastosis virus reverse transcriptase (5 units, RT, Promega) using random hexamers (Invitrogen) for 1 h at 42°C. The cDNA was subsequently amplified using standard PCR conditions as described (Lukashevich et al. 2002). RESULTS Viremia was detected 4-6 days after inoculation with LCMV-WE - The viral infection of rhesus macaques inoculated with 103 pfu LCMV-WE i.v. was confirmed by RT-PCR using specific primers for the envelope glycoprotein as described (Lukashevich et al. 2002). Since the infection data for these animals has been published (Lukashevich et al. 2004), we will summarize here that viral sequences were detected in plasma as early as four days after inoculation and virus was detected by plaque assay of plasma at 4-6 days after the inoculation. As our group and others have previously described, the only early evidence of infection in the rhesus model, besides circulating virus, is a dramatic increase of liver enzymes (Lukashevich et al. 2003). Other signs of the viral disease are barely detectable before 8-9 days after inoculation. Circulating NK cells decrease soon after LCMV infection - According to flow cytometric analysis and manual differential cell counts, there were no dramatic or consistent changes in the major circulating blood cell populations (Fig. 1A-C). Absolute numbers of CD3+CD4+, CD3+CD8+ and CD20+ cells decreased in only two of the monkeys (2nd and 6th), while the values of these subsets did not change significantly for the other monkeys (Fig. 1D-G). On the other hand, the number of cells that were CD3-CD8+, also recognized as the phenotype for NK cells (Dykhuizen et al. 1998, 2000), increased slightly during the first day after infection, though the absolute numbers then decreased consistently in all the monkeys tested > 2 days after inoculation (Fig. 1H). Specific examples of decreasing NK cells (CD3-CD8+) are seen by flow cytometry (Fig 2A, B). The absolute numbers and average of NK cells are also shown for two groups of samples: those taken before inoculation and those taken on the day of euthanasia (between days 2 and 7) (Fig. 2C). The t-test establishes a statistically significant probability (p = 0.022) that the two groups of samples are different. As a control for the analysis of lymphocyte subsets in LCMV-WE-infected monkeys, we also examined two LCMV-ARM-infected monkeys that never experienced disease (i.e., no rise in liver enzymes or body temperature) (Rodas et al. 2004). These animals were analysed before intravenous infection with 103 pfu (Arm iv3) or intragastric infection with 108 pfu (Arm ig8) of LCMV-ARM strain, two weeks following infection, before and two weeks after a lethal challenge with 103 pfu of LCMV-WE i.v. given eight weeks after the first inoculation and then, after surviving the LCMV-WE challenge unscathed, both were sampled before a second challenge with LCMV-WE. A week after the second challenge circulating NK cell numbers increased rather than decreased (absolute and percentage). Similar to the LCMV-WE-infected animals, they displayed a 16-30% decrease in circulating NK cells after infection (Fig. 3). This was less than the mean decrease seen with the LCMV-WE-infected monkeys, but the difference was not statistically significant because so few animals were examined. By three weeks after infection, circulating NK cells had been restored to the numbers seen before infection. NK and NKT cell subsets - To better characterise the NK cells, several markers previously reported for this cell subtype were employed. Using monoclonal antibodies for CD16 and CD56, it appeared that the numbers of CD3-CD16+ cells (i.e., the majority of the CD3-CD8+ NK cells) disappeared from the circulation during the course of infection. The CD3-CD56+ cells are a barely detectable sub-population of the CD3-CD8+ NK cells (Fig. 4A-C). Absolute numbers of cells are presented in Fig. 4, but a similar picture is obtained by looking at percentages of circulating PBMCs (not shown). In the overall picture for NK cells, whether we consider the CD3-CD8+ cells or the majority of these that are CD3-CD16+, we see that they represented 4-15% of PBMC before infection and they became 2-7% of PBMC after two days of infection. NKT cells were also observed in the LCMV-infected monkeys. The numbers of circulating NKT cells (CD3+CD16+ and CD3+CD56+) were even lower than the numbers of NK cells (note the different scales of CD3+ and CD3- cells: 100 compared to 350 on the "y" axis on Fig. 4D-F versus Fig. 4A-C), representing only 1-2% of PBMCs. According to Judy Thomas, who has been working with rhesus NK cells, the proportion of circulating NKT cells is lower than 0.23 ± -0.12 (personal communication). We saw a slight increase in the NKT population (both percentage and absolute number) during the course of infection, but this was not statistically significant (Fig. 4D-F). Since CD3+CD16+ cells often include γδ T cells (Eberl et al. 2005), we also examined that population using flow cytometry. Vγ9 T cells, which represent approximately 80% of the γδ T cells in primate peripheral blood, exhibited a similar pattern of reduction in the percentage and absolute numbers as the NK cells (CD3-CD8+) after the second day of infection (Fig. 4G). Some of the NK cell decrease may be attributable to cell death - To determine whether the decrease in circulating NK cells could be attributable to virus-mediated cell death, we incubated monkey or human PBMCs with virus (LCMV-ARM or LCMV-WE) at a multiplicity of one pfu/cell and looked for apoptosis using 7-AAD staining. The reduction in absolute numbers of NK cells in the human PBMC samples (Fig. 5E) is similar to the reduction of percentages seen in the monkey and human samples (Fig. 5A-D). Absolute cell counts were determined by counting cells in triplicate cultures and then using the percentage of particular markers determined by flow cytometry. The apoptosis identified via 7-AAD staining was negligible (< 2%) in both monkey and human samples (not shown). Although the number of cells undergoing apoptosis was negligible, absolute NK cell numbers were reduced almost 30% over 72 h of cell culture, indicating significant cell death due to culture alone (Fig. 5E) (trypan blue staining). The WE-infected cultures experienced approximately 7% more cell death than the uninfected and ARM-infected cultures, raising the possibility that virus-mediated cell death could be contributing (less than 10%) to reductions in NK cells. The reduction in circulating NK cells is not likely to be due to cell death, but the reduction in Vγ9 TCR-positive cells may be partially explained by apoptosis. Apoptosis was not a significant cause of the NK cell decrease, but it did seem to affect the γδ T cell population. Two of the monkeys depicted in Fig. 1, which showed typical reductions in NK cells (CD3-CD8+) and γδ T cells, were also monitored for apoptosis of circulating NK cells. Apoptosis was measured via 7-AAD staining and was insignificant (< 5%) in the circulating NK cell population during the course of infection (Fig. 6). The reduction of Vγ9 T cells, on the other hand, was accompanied by 10-50% 7-AAD staining, indicating rather high levels of apoptotic cell death. DISCUSSION The virulence of a pathogen is often determined by innate responses of the host soon after infection. To understand the events leading to disease, we described changes in circulating blood cells occurring soon after infection. In 10 rhesus macaques, we observed a mean 55% reduction in circulating NK cells from 2-7 days after lethal LCMV-WE infection. The reduction in NK cells, which ranged from 20-70%, encompassed the NK cell drop of 16-30% observed in two monkeys infected with the non-pathogenic strain, LCMV-ARM. Whereas the monkeys infected with the benign virus (ARM) recovered their circulating NK cells within two weeks, lethally infected monkeys (WE) generally succumbed by day 12 and showed no recovery of circulating NK cells. Another 20-30% drop in NK cells was observed when the LCMV-ARM-infected animals were given a lethal challenge with LCMV-WE, but when they were given a second challenge, NK numbers were not reduced. Thus, either the second challenge failed to elicit an inductive signal or the NK cells had become unresponsive to the inductive signal. Although our study failed to show a significant difference between the benign and virulent infections, it is possible that a study with more time points could detect kinetic differences in NK loss differentiating these two infections. Since ARM-infection immunises monkeys from lethal LCMV-WE challenge (Danes et al. 1963, Peters et al. 1987, Rodas et al. 2004), it is notable that a drop in NK cells is seen during a WE challenge infection in the absence of pathogenesis. Murine infections with LCMV have provided a large body of information on innate immune responses after viral inoculation. Interferon type I (or IFN α/β) is produced by infected cells soon after LCMV infection of the mouse and it then activates NK cells to proliferate and carry out their cytotoxic role (Biron et al. 1996, Nguyen et al. 2002). Type I IFN inhibits the expression of IL-12 (Biron et al. 1996, Cousens et al. 1999, Pien et al. 2000) and in the absence of IL-12, type II interferon (IFN-γ) increases in peripheral organs without inhibition (Cousens et al. 1997, 1999, Pien et al. 2000). Again, in the mouse model during the acute phase of LCMV infection, NK cells increase in the peripheral blood by 10% over three days and NK cell margination results in a > 30% increase in the liver (McIntyre & Welsh 1986, Biron et al. 2002). Murine NK cells have been pheno typically characterised as NK 1.1+TCR-, CD3-, CD8+, asialo+ and CD16+; they are known to stimulate the induction of T cell-mediated immunity by secreting IFN-γ and they also cause some cell death in the liver (Liu et al. 2000). In our monkey study, where NK cells are primarily CD3-CD16+, we did not investigate the migration of NK cells to peripheral organs, but in other published studies, we have demonstrated the presence of unidentified cellular infiltrates in the liver and lymphoid tissues (Lukashevich et al. 2002, 2003, 2004). Here, we demonstrated in cell culture that virus-mediated cell death could not satisfactorily explain the reduction in circulating NK cells. Although NK cells account for approximately 5% of circulating PBMCs, the high interest in NKT cells caused us to monitor changes in that population as well, even though they account for less than 1% of circulating PBMCs in primates (Koyasu 1994). NKT is a subpopulation of T cells with common markers for NK cells, but higher heterogeneity. NKT cells have been identified in the mouse as either double negative for CD4 and CD8 or as positive for CD4 with a very limited diversity of α/β TCR, most commonly Vα14 Vβ281 (Emoto & Kaufmann 2003). In human beings, the NKT TCR is Vα24, which cross-reacts with the rhesus monkey cell determinant for NKT (Gansuvd et al. 2003). In rhesus macaques, 80% of Vα24TCR+ cells from the spleen also express CD56 (Gansuvd et al. 2003). Although it has been accepted that rhesus macaque NKTs are CD56+, there is some controversy about the presence of CD16 on their surface (Gansuvd et al. 2003), so we used both the CD16 and CD56 as markers. Unlike the CD3- NK cells, the NKTs in our monkey studies seemed unaffected or even a little increased by day 7 of the study. A rise in NKTs, concomitant with a drop in NK cells has been reported in murine studies (Eberl & MacDonald 2000, Hobbs et al. 2001) and has been interpreted to indicate complementary functions for the two cell types. Specifically, murine infection with both LCMV-ARM and LCMV-WE resulted in a 30% increase in NK cells in the liver by day 3 (Pien et al. 2000), whereas LCMV-ARM infection resulted in an apoptotic reduction of NKT cells in the liver, where this population is commonly most abundant (Hobbs et al. 2001). Blood cells before and after infection were examined for alterations in apoptosis using 7-AAD, a fluorescent DNA-binding agent that preferentially labels cells with reduced plasma membrane integrity (Lecoeur et al. 1997). No significant increases in the levels of apoptosis were observed in NK cells derived from fresh PBMCs or from cells cultured in the presence of virus, although non-apoptotic cell death of about 30% (of infected or non-infected cells equally) was observed in these cultures. In the murine system, apoptosis was only seen in NKT cells that were already residing in the periphery (Hobbs et al. 2001). Since the NKT cells were not susceptible to LCMV infection, they were most likely eliminated by a bystander effect such as that described during HIV infection (Finkel et al. 1995, Abbate et al. 2000, Yang et al. 2003). One likely mechanism is that abundant IFNα/β in peripheral organs increases double-strand RNA-dependent protein kinase in uninfected cells that induces apoptosis (Biron 1999, Biron et al. 2002). Ebola infection of non-human primates resulted in apoptosis of NK cells, as shown by TUNEL assays of NK cells in PBMCs, liver, spleen and lymph nodes (Geisbert et al. 2000, 2003a, b, Reed et al. 2004). Although we saw little evidence of apoptosis in LCMV-infected monkey PBMCs, we could not rule out an almost 10% non-apoptotic reduction in NK cell numbers cultured in the presence of LCMV-WE. Since virus does not infect NK cells, the virus-mediated loss of NK was most likely due to indirect effects or to cell-surface signalling by viral particles. Therefore, the reduction in circulating NK cells in the primate model can most likely be attributed to margination of NK cells to peripheral organs with a lesser proportion due to virus-mediated cell death. Evidence that NK cells have little influence on virus loads in murine LCMV infections first came from the Welsh laboratory. They have reported that (i) virus titers were unperturbed during the early stages of infection in mice depleted of NK cells by cyclophosphamide (Biron et al. 1996); (ii) Beige mice (NK deficient) synthesized normal levels of virus (Roder & Duwe 1979); (iii) adoptive transfer of NK cells into 5-day-old mice with low NK cell activity did not confer LCMV resistance even though it conferred resistance to murine CMV (Bukowski et al. 1985) and (iv) depletion of NK cells (using antibody to asialo-GM1) had no effect on the synthesis of LCMV-WE or LCMV-ARM in acute or persistent infections (Bukowski et al. 1983, Welsh et al. 1984). Although NK cells have little impact on LCMV titers, they are purported to play a role in controlling viral dissemination (Welsh et al. 1984) and in modulating T cell responses during infection (Su et al. 2001). NK cells produce IFNγ, which induces the expression of major histocompatibility complex class I on antigen-presenting cells, thereby stimulating CD8+ lymphocytes that play an important role in controlling LCMV infection (Oldstone 2002). If NK cells do impact virus dissemination, one might predict that intravenous infections that are systemically disseminated and rapidly established in peripheral organs might escape the effects of NK cells, whereas more slowly disseminated infections, such as mucosal or subcutaneous infections, might be attenuated by NK cells. Similar to NK cells, γδ T cells have little effect on murine LCMV infection and CTL-mediated clearance (Pien et al. 2002, Eberl et al. 2005). γδ T cells have been implicated in immunosurveillance of tumours and infections via cytokine secretion and cytolysis (Poccia et al. 2002, Jacobs et al. 2005). In contrast with the absence of virus-mediated cell death in cultured NK cells, γδ T cells display a much higher sensitivity to virus exposure in culture. This has been described for exposure with HIV and is thought to be a direct activation of apoptotic signals by the virus (Cairo et al. 2005). Thus, the decrease in circulating γδ T cells may constitute a viral strategy for eliminating a potential suppressor of viral infection.Recently, Baize et al. (2009) have reported a NK cell decrease early in Lassa virus-infected cynomolgous macaques like we described in LCMV-infected rhesus macaques. ACKNOWLEDGEMENTS To excellent animal care facilities and pathology support from Harry Davis. REFERENCES
Copyright 2009 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc09134f3.jpg] [oc09134f6.jpg] [oc09134f1.jpg] [oc09134f4.jpg] [oc09134f2.jpg] [oc09134f5.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}