 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 104, No. 4, July, 2009, pp. 636-643 ARTICLES Molecular approaches for eco-epidemiological studies of Paracoccidioides brasiliensis Virgínia Bodelão Richini-Pereira; Sandra de Moraes Gimenes Bosco; Raquel Cordeiro Theodoro; Severino Assis da Graça Macoris; Eduardo Bagagli+ Departamento de
Microbiologia e Imunologia, Instituto de Biociências, Universidade Estadual
Paulista, Distrito de Rubião Júnior s/n, 18618-000 Botucatu, SP,
Brasil Financial support: Fapesp (05/56771-9 and 06/03597-4) Received 15 November
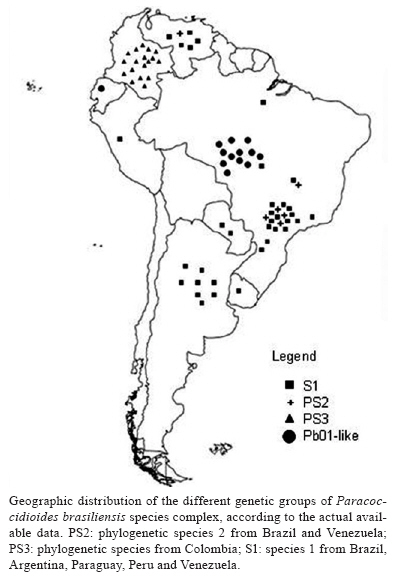
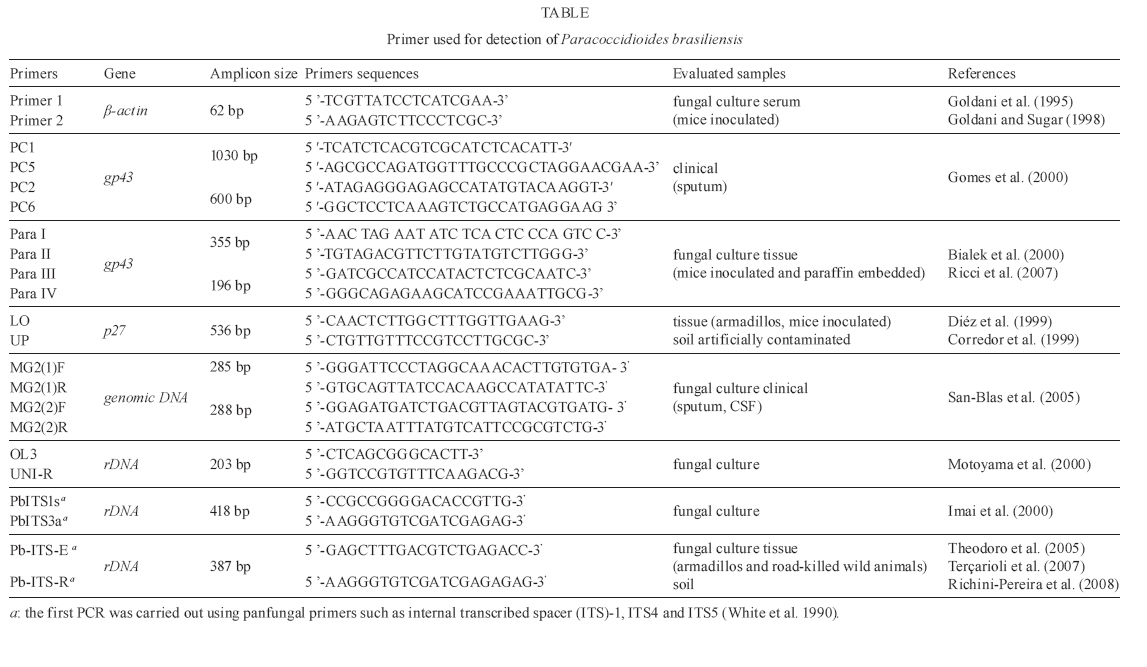
2008 Code Number: oc09143 ABSTRACT Medical mycology has greatly benefited from the introduction of molecular techniques. New knowledge on molecular genetics has provided both theoretical and practical frameworks, permitting important advances in our understanding of several aspects of pathogenic fungi. Considering Paracoccidioides brasiliensis in particular, important eco-epidemiological aspects, such as environmental distribution and new hosts were clarified through molecular approaches. These methodologies also contributed to a better understanding about the genetic variability of this pathogen; thus, P. brasiliensis is now assumed to represent a species complex. The present review focuses on some recent findings about the current taxonomic status of P. brasiliensis, its phylogenetic and speciation processes, as well as on some practical applications for the molecular detection of this pathogen in environmental and clinical materials. Key words: Paracoccidioides brasiliensis - eco-epidemiology - PCR Paracoccidioidomycosis (PCM) is the most important and prevalent systemic mycosis in Latin America, occurring mainly in Brazil, Colombia and Venezuela (Wanke & Londero 1994). It is caused by Paracoccidioides brasiliensis, a thermally dimorphic fungus that grows as a yeast-like structure in host tissues or when cultured at 35-37°C and as a mycelium under saprobic conditions or when cultured at rt, 18-23°C (Lacaz 1994). PCM-infection is believed to be acquired through the inhalation of conidia present in the environment, which is similar to other systemic mycoses also caused by dimorphic fungi and in which the lung is the main compromised organ (Gonzales-Ochoa 1956, Bustamante et al. 1985). Although PCM is considered an endemic disease in many regions, there are few reports in the literature about the isolation of this pathogen from the environment, which has made ecological studies on P. brasiliensis difficult for mycologists (Shome & Batista 1963, Negroni 1966, Albornoz 1971, Restrepo 1985, Silva-Vergara et al. 1998, Franco et al. 2000). There have been references to P. brasiliensis isolation from bat and penguin faeces and dog food. However, these were casual remarks with no reproducibility (Grose & Tamsitt 1965, Gezuele 1989, Ferreira et al. 1990). Besides, the lack of outbreaks and the prolonged latency period of this disease, together with human migration, allow the exact infection source to remain unknown (Restrepo 1985). An important clue for ecological studies on P. brasiliensis was the finding of naturally infected nine-banded armadillo (Dasypus novemcinctus) in endemic areas (Naiff et al. 1986, Bagagli et al. 1998, 2003, Corredor et al. 1999, Silva-Vergara & Martínez 1999). The fungus was also isolated from another armadillo species, Cabassous centralis, reinforcing that armadillos are in constant contact with the pathogen in the environment (Corredor et al. 2005). The systematic recovery of P. brasiliensis from armadillo tissues has demonstrated the importance of this animal in PCM endemic areas, helping locate hot spots of fungal occurrence in some environments and suggested valuable insights about its evolutionary aspects (Bagagli et al. 2006, 2008). The environment represented by the armadillo burrow and its surroundings, associated with biotic and abiotic features, may contribute to the development of the fungus saprobic stage in nature, as already demonstrated by Terçarioli et al. (2007). For a long time, armadillos had been the only wild animal species in which fungus recovery from tissue cultures was possible, although many attempts have been made in other animal species from PCM endemic areas. Recently, other wild mammals such as guinea pig (Cavia aperea), porcupine (Sphiggurus spinosus), raccoon (Procyon cancrivorus) and grison (Gallictis vittata) have been recognized through molecular methods as new hosts for this infection (Richini-Pereira et al. 2008). Learning about the natural reservoirs of this pathogenic fungus may contribute to mapping of endemic areas and to a better understanding of its eco-epidemiological features. However, some questions still need to be answered, such as: does the fungus need some special substrate or a specific period to grow? Which are the most appropriate environmental conditions, climates and soil types? Which host-parasite relationship does the fungus establish? Is there another infection route, besides the airborne route? Medical mycology has extensively benefited from the great development of molecular biology in the latest years. Thus, the aim of this review is to present some recent advances in the understanding of the eco-epidemiology of P. brasiliensis and related species through molecular approaches. Molecular biology has demonstrated that P. brasi-liensis belongs to a peculiar Ascomycota family Morphological and molecular findings have suggested that the main pathogenic fungi causing systemic mycosis had been classified into Ajellomycethaceae, a new family of vertebrate-associated Onygenales, which includes fungi of the genera Histoplasma, Blastomyces, Emmonsia and Paracoccidioides (Untereiner et al. 2004). The correct taxonomic position of these fungi has opened new possibilities for their study and understanding of their eco-epidemiological relationships with their respective hosts. This fungal group (Onygenales, Onygenaceae sensu lato) presents several common mycological and ecological features such as dimorphism, arthroconidia production and restricted geographic distribution. In addition, the natural affinity of some of its members, such as Blastomyces dermatitidis and Histoplasma capsulatum, for animal product derivatives or remnants like faeces and uric acid is well documented (Baumgardner & Paretsky 1999, Restrepo 2000, Untereiner et al. 2004). Several lines of evidence indicate that P. brasiliensis is phylogenetically closer to Lacazia loboi than to Blastomyces dermatididis, Emmonsia parva and H. capsulatum (Bialek et al. 2000, Herr et al. 2001). It must be emphasized that L. loboi could not yet be cultured on standard media and is therefore considered an obligate parasite (Taborda et al. 1999, Herr et al. 2001). This suggests a possible tendency for reduction or extinction of the fungus saprobic form in nature. Lacaziosis, the mycosis caused by L. loboi, is acquired through traumatic route and evolves typically as a chronic subcutaneous infection. It occurs in a restricted region of the Brazilian Amazon, Surinam and some other countries in northern South America and Central America. It has also been observed in some wild aquatic mammals, especially Atlantic dolphins (Herr et al. 2001). It must be considered that this phylogenetic group (Onygenales) includes the species E. parva and Emmonsia crescens that cause adiaspiromycosis, a localized lung infection of cosmopolitan distribution, both in wild animals and humans (Hubalék et al. 1998). Some aspects of their saprobic and parasitic stages have led Emmonsia species to be considered close relatives of dimorphic fungi (Emmons & Ashburn 1942, Sigler 1996, Peterson & Sigler 1998). Their occurrence has been related to the agroecosystem and their highest abundance was observed in plant remnants and rodent burrows (Hubalék et al. 1998). The presence of this fungus has been reported in domestic and wild rodents (Zlatanov & Genov 1975, Hubalék et al. 1998), carnivores (Krivanec et al. 1980) and armadillos (Santos 1999). Studies have also indicated that the different Onygenaceae (sensu lato) species may have originated in the Americas around 3-20 million years ago (Fisher et al. 2000) and some of the most important ones, such as Histoplasma and Paracoccidioides species, had certainly evolved in South America prior to the Panamanian Isthmus connection (Kasuga et al. 1999, 2003). Molecular phylogenetics indicates that P. brasiliensis is a species complex Like all living organisms, P. brasiliensis has been shown to present a significant diversity, which has been well documented by different mycological, antigenic and virulence studies, as well as by molecular techniques (Franco et al. 1996, Calcagno et al. 1998, Sano et al. 1999, Hebeler-Barbosa et al. 2003a, b, Macoris et al. 2006, Puccia et al. 2008). Instead of a unique species, P. brasiliensis appears to contain several different cryptic species, as observed by phylogenetic studies using Multi-Locus Sequence Type (MLST) (Matute et al. 2006a, Carrero et al. 2008), microsatellite analysis (Matute et al. 2006b) and prp8 intein sequencing (Theodoro et al. 2008a). Unlike the morphological and biological concepts of species, which are rarely applicable to separate fungal species, the Phylogenetic Species Concept consider a species as a cluster of individuals that presents exclusive apomorphic characters and is genetically isolated from other species (i.e. individuals from different species are not exchanging alleles). According to this concept, the Phylogenetic Species Recognition (PSR) by MLST uses several nuclear genes and detects the species limits through genealogical concordance (the clade must be present in the majority of the single-locus genealogies) with high bootstrap and posterior probability values supporting the clades (Taylor et al. 2000). Applying the PSR approach, Matute et al. (2006a, b) analysed eight regions of five nuclear coding genes: chitin synthase, β-glucan synthase, α-tubulin, adenyl ribosylation factor and PbGP43 and also performed microsatellite analysis that revealed three distinct, previously unrecognised species: S1 (species 1 from Brazil, Argentina, Paraguay, Peru and Venezuela), PS2 (phylogenetic species 2 from Brazil and Venezuela) and PS3 (phylogenetic species from Colombia). They also observed that S1 and PS2 were both recombining sexual species, whereas PS3 was shown to be clonal. It is interesting to note that S1 and PS2 are sympatric but reproductively isolated, suggesting an intrinsic biological barrier to reproduction. The divergence time between PS2-PS3 was estimated as 8.04-8.37 mya through chitin synthase gene analysis (Matute et al. 2007). Those authors suggested that the speciation of PS3, geographically restricted to Colombia, could be attributed to dispersal, leading to genetic isolation of PS3 from S1 (allopatric speciation). The other speciation event that originated PS2 is still poorly understood. S1 and PS2 may be the result of sympatric speciation, the initial step of which is the existence of polymorphism, much more abundant in recombining species such as PS2 and S1. Therefore, two forms of a single species can be adapted to different conditions in their niche, leading to negatively selected interbreeding due to the low adaptive value of the hybrids (Coyne & Orr 2004). Recently, more P. brasiliensis isolates from the Central Region of Brazil were included in a new MLST analysis and showed a significant genetic divergence when compared with other S1, PS2 and PS3 isolates (Carrero et al. 2008, Teixeira 2008). This cluster of isolates was named Pb01-like, since the first studies that revealed such divergence were carried out with Pb01 isolate. Phylogenetic analyses with several nuclear encoding regions such as GP43, CH4, Actin, ODC, URA3, CHS2, FKS1, HSP70, Hydrophobin, Kex, Catalase A, Catalase P, Formamidase and Glyoxalase showed a significative genetic distance between Pb01 and the remaining genetic groups (Carrero et al. 2008). Pb01-like isolates form a well-supported clade for several nuclear coding genes, suggesting the existence of one more cryptic species of P. brasiliensis (Carrero et al. 2008, Teixeira 2008). The divergence time between Pb01-like and the remaining clades was estimated as 20 mya, a long evolutionary time (Teixeira 2008). However, it is not clear yet which factors (geographic, biotic and abiotic) could have contributed to the genetic isolation of this group right in the Central Region of Brazil. This Pb01 isolate, which has been the subject of genome and transcriptome projects (Felipe et al. 2005), also substantially differs from the others in the hsp70 gene, which encodes a conserved heat shock protein. Four large insertions: one of four nucleotides, one of 16, a third one of 23 and another one of 21 (the last corresponds to a CT microsatellite), were observed in the hsp70 gene of Pb01, relative to P. brasiliensis isolates from the other species (Teixeira et al. 2005, Theodoro et al. 2008b). An additional molecular marker, the PRP8 intein, was used to recognize P. brasiliensis isolates of the different genetic groups (Theodoro et al 2008a). Inteins are coding sequences that are transcribed and translated with flanking sequences (exeins). After translation, inteins are excised by an autocatalytic process; then, the host protein assumes its normal conformation and can develop its function (Cooper & Stevens 1995). These parasitic genes have been found in several vital proteins in all three domains of life and are widespread in fungi. The PRP8 intein is known to occur in important fungal pathogens such as Cryptococcus neoformans (varieties grubii and neoformans), Cryptococcus gattii, H. capsulatum and P. brasiliensis (Butler & Poulter 2005, Butler et al. 2006). There are two intein types, mini-inteins and full-length inteins, both of which have a splicing domain. Full-length inteins additionally present an endonuclease domain that can play a homing function (Homing Endonuclease Gene or HEG), which makes intein a mobile genetic element, resulting in both the occupation of empty alleles and the duplication of parasitic genetic element (Liu 2000). These large inteins are expected to have more sequence variation in the endonuclease domain than in the splicing domain due to a more relaxed selection, especially if the HEG is no longer active (Gorgaten & Hilario 2006), which makes inteins a promising source of phylogenetic information. In this assay, the PRP8 inteins of 22 P. brasiliensis isolates belonging to all four previously recognized species were sequenced. The phylogenetic analysis clearly separated the isolates from the four species and revealed a significant difference between Pb01-like and the remaining species (Theodoro et al. 2008a). All evaluated P. brasiliensis isolates presented a full-length intein in their prp8 gene. The HEG domain of PRP8 intein from P. brasiliensis appears to be inactive due to a substitution in the second aspartic acid residue, which is indispensable to its functionality. Although the degree of polymorphism in the PRP8 intein from P. brasiliensis was not as high as that in the commonly used nuclear-coding sequences, its sequence contains sufficient phylogenetic information to separate cryptic species of P. brasiliensis, constituting therefore a reliable additional molecular marker for this pathogen. Since distinct cryptic species have been recognized through molecular techniques, it becomes easier to detect other biological features that could be associated with the different genetic groups. Theodoro et al. (2008b) detected some microscopic features of S1 and PS2 isolates that could be important candidates for a morphological differentiation between these two species. The isolates T10 and Bt84, representing PS2, seemed to have elongated yeast cells and presented a slower Mycelia-to-yeast transition when compared with the other isolates. In addition, Terçarioli et al. (2007) studied fungal growth and conidia production on Soil Extract Agar and observed that most S1 group isolates produced large quantities of conidia; this was not observed in PS2 group isolates. Both S1 and PS2 genetic groups are sympatric in the same endemic area; however, S1 is curiously much more frequent in both patients and armadillos, probably due to its higher conidium production (Bagagli et al. 2008). As a large number of isolates from different areas are molecularly characterized, the geographical distribution of such strains becomes clearer. Thus, S1 appears to be widely distributed, sympatrically occurring with PS2 in several endemic areas; the PS3 group is centralized in Colombia; and the Pb01-like group, in the Central Region of Brazil. The geographic distribution of the different genetic groups of the P. brasiliensis species complex is summarized in Figure, according to the currently available data. Since prophylactic or treatment measures for PCM must include all genotypes and/or phenotypes of P. brasi-liensis, the discovery of cryptic species increased the importance of comparative studies aiming to detect relevant phenotypic differences among species. These studies may be of help in correct diagnosis and treatment of PCM. Pathogen detection - clinical and environmental sources PCM diagnosis is usually made by microscopic detection of characteristic multibudding yeast cells and/or by isolation of fungal cultures from clinical specimens, histopathology of biopsied tissue and serological tests, especially including gp43, a reference P. brasiliensis antigen (Lacaz 1994). Each of these procedures has advantages and disadvantages. Although a definitive diagnosis can be made through fungal culture, it must be considered that such procedure involves high contamination risk, long incubation period and low sensitivity due to the low amounts of fungus in some clinical samples (Salina et al. 1998, Sano et al. 2001). In histological sections, the etiological agent might be undetected in some pauci-cellular samples or confused with other dimorphic fungi (Bialek et al. 2000). Several serology techniques using different P. brasiliensis antigen types have been employed for both diagnosis and monitoring therapy of patients; however, problems such as anergy and cross-reactivity as well as the specificity and sensitivity of the technique must be considered (Albuquerque et al. 2005). Recent advances in molecular biology, mainly based on the PCR, provide powerful tools for the detection, identification and typing of different pathogen groups. The main genomic regions employed for primer designs for use in both clinical and environmental samples of P. brasiliensis are listed in Table. Goldani et al. (1995) designed specific primers based on the β-actin gene which amplified a 62bp-fragment not detected in other fungi such as H. capsulatum, B. dermatitidis, C. neoformans, Candida albicans, Aspergillus fumigatus, Saccharomyces cerevisiae and Pneumocystis carinii. Later, Goldani and Sugar (1998) employed this same set of primers to detect DNA of P. brasiliensis in blood from five experimentally infected mice and compared this result with culture. They concluded that the molecular approach was more sensitive rather than culture (100% vs. 40%, respectively) and suggest further evaluation of such approach in different clinical forms of PCM to assess the true value of this diagnostic method. PCR assays targeting the gp43 gene (PbGP43) are widely used for the molecular detection of P. brasiliensis DNA in several types of samples Cisalpino et al. (1996) cloned and characterized the entire coding region of the gp43 gene, which comprises two exons interrupted by a 78bp-intron. After this study, several sets of primers were designed and employed for molecular diagnosis. Gomes et al. (2000) combined five primer pairs and suggested a PCR using the PC2-PC6 primer pair for direct amplification from clinical material (sputum) or for nested-PCR following amplification with the PC1-PC5 primer set. In an experimental model infected with yeast cells of H. capsulatum and P. brasiliensis, Bialek et al. (2000) employed a nested-PCR, also based on gp43, with high sensitivity and specificity for the sets of primers para I/II and para III/IV. Using these same sets of primers, Ricci et al. (2007) demonstrated 30% positivity in pathogen detection in biopsies from PCM-patients. Those authors emphasized that this low positivity might be due to the procedures used for fixation, paraffin embedding and storage of the material, which favoured DNA degradation and to the use of primers for regions of gp43 that have proven to be polymorphic. This fact may have interfered with the annealing of the primers employed (Morais et al. 2000, San-Blas et al. 2002). Since gp43 is a highly polymorphic gene that clearly separates the four cryptic species of P. brasiliensis (Matute et al. 2006a, Carrero et al. 2008, Teixeira 2008), PCR based on this genomic region is now indicated not only for pathogen detection, but also for genotyping of clinical samples. By using this strategy, Ricci et al. (2008) evaluated and distinguished between S1-PS2 from paraffin-embedded tissue. Besides the gp43 glycoprotein, a 27-kDa antigen protein was cloned, sequenced and characterized by McEwen et al. (1996). The use of the LO and UP primer combination allowed the detection of P. brasiliensis DNA in tissue samples of armadillos and inoculated mice as well as in artificially contaminated soil (Corredor et al. 1999, Díez et al. 1999). Internal transcribed spacer (ITS) regions, including the 5.8S ribosomal RNA of P. brasiliensis, are also employed for molecular detection and distinction from other fungal species. It is known that ribosomal DNA (rDNA) genes are present in all microorganisms and have regions that accumulate mutations both at slow (28S, 5.8S and 18S) and high (ITS1 and ITS2) rates over time. The first regions provide a molecular basis for establishing phylogenetic relationships among taxonomic levels above genus and the second regions are useful for the separation between genera and species. Besides, this target provides higher sensitivity to PCR due to its multiple copies per genome (more than 100 copies) (White et al. 1990, Iwen et al. 2002). This region has been considered the main candidate for a Barcoding system of fungal identification (Buckley 2008). PCR of rDNA (5.8S-ITS) can be amplified by panfungal primers and the sensitivity and specificity of this reaction may be significantly improved by Nested-PCR with species-specific inner primers. Using this method, Imai et al. (2000) designed a set of primers, PbITS1s and PbITS3a, for nested-PCR and identified 29 P. brasiliensis strains by means of a specific 418bp-fragment, which was not detected in A. fumigatus, B. dermatididis, C. albicans, C. neoformans, H. capsulatum, or Penicillium marneffei. Motoyama et al. (2000) employed the OL3 and UNI-R primer combination for PCR and obtained a 203bp-fragment when P. brasiliensis (Pb01) DNA was used as the template. These primers were capable of discriminating between P. brasiliensis and H. capsulatum. A nested-PCR, also using species-specific inner primers (PbITSE and PbITSR) derived from ITS-5.8S rDNA, was developed for detection of P. brasiliensis in soil (Theodoro et al. 2005, Terçarioli et al. 2007). This technique also proved to be useful for fungal detection in road-killed wild animals (Richini-Pereira et al. 2008). As P. brasiliensis actually consists of a species complex with distinct genetic groups, it must be emphasized that all molecular protocols should be re-evaluated both in silico and experimentally to detect any genotype that cause PCM in endemic areas. REFERENCES
Copyright 2009 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc09143f1.jpg] [oc09143t1.jpg] |
| |||||||||

{kind=link}
{kind=link}