 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 104, No. 5, August, 2009, pp. 683-688 ARTICLES Effect of Solanum nudum steroids on uninfected and Plasmodium falciparum-infected erythrocytes Mary Luz LópezI; Silvia BlairI; Jairo SáezII; Cesar SeguraI, + IGrupo
Malaria, Sede de Investigación Universitaria, Universidad de Antioquia,
Calle 62 52-59, Laboratory 610, Medellín, Colombia Financial support: COLCIENCIAS (MLL) (1115-05-13667, 1115-05-16877), Universidad de Antioquia, Medellín-Colombia Received 28 November
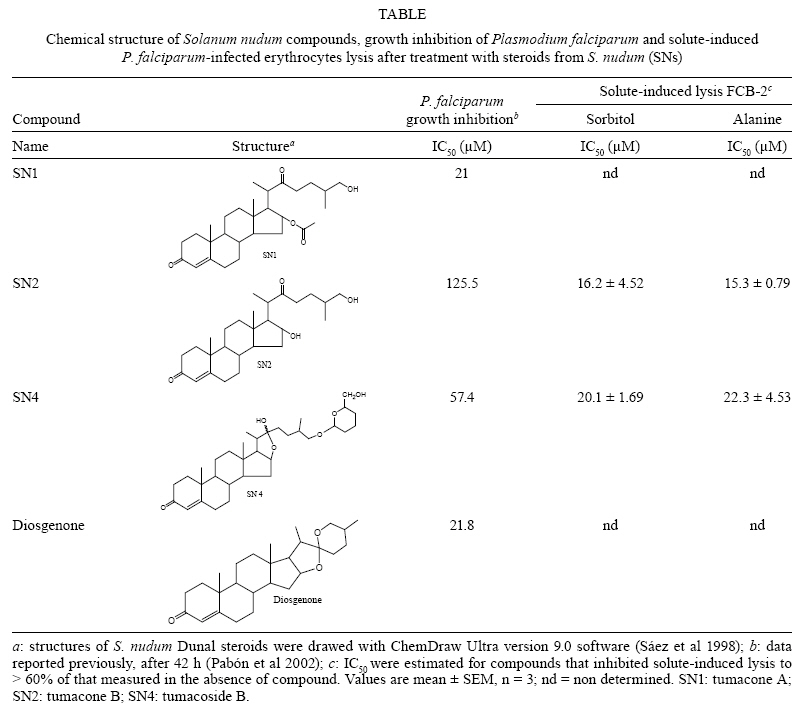
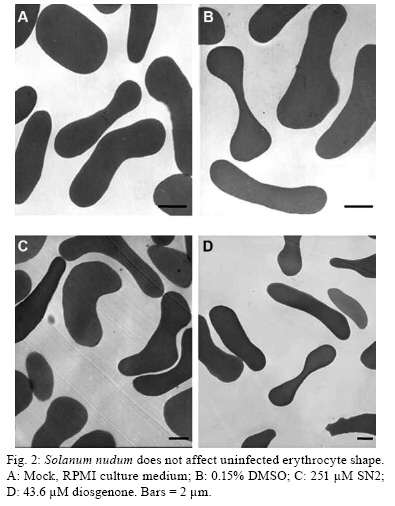
2008 Code Number: oc09154 ABSTRACT Steroids from Solanum nudum (SNs) have demonstrated antiplasmodial activity against erythrocytic stages of the Plasmodium falciparum strain FCB-2. It is well known that steroids can alter the membrane function of erythrocytes. Thus, we assessed alterations in the membranes of uninfected red blood cells, the parasite invasiveness and the solute-induced lysis of parasitised red blood cells (pRBCs). induced by SNs. We found that most merozoites were unable to invade SN-treated erythrocytes. However, transmission electron microscopy revealed no effect on the morphology of uninfected erythrocytes treated with either SN2 or diosgenone and neither SN induced haemolysis of uninfected erythrocytes. SN2 and SN4 inhibited isosmotic sorbitol and alanine-induced haemolysis of pRBCs. In contrast, diosgenone and SN1 did not inhibit solute-induced haemolysis. The inhibition of solute-induced lysis of parasitised erythrocytes by SN2 and SN4 suggest an action of these SNs on new permeability pathways of pRBCs. Key words: erythrocyte - Solanum nudum - Plasmodium falciparum - new permeability pathways - ultrastructure - malaria Malaria is a significant public health problem in the developing countries. In humans, the most severe form of malaria is caused by Plasmodium falciparum and at least one-third of the world's population is at risk of infection, with over 300 million people developing clinical disease each year and at least two million deaths (Snow et al. 2005). The emergence of drug-resistant malaria parasites and the lack of new antimalarial drugs of proven efficacy have contributed to a global increase in the number of malaria cases. Therefore, research into new antimalarial drugs active against multidrug-resistant P. falciparum is urgently needed. P. falciparum-infected erythrocytes display several dramatic morphological changes that affect membrane rigidity, increased cell adhesiveness, permeability and susceptibility to haemolysis (Kirk 2001, Staines et al. 2004b). The changes in parasitised red blood cells (pRBCs) are intimately connected to Plasmodium biology and are involved in nutrient acquisition through new permeability pathways (NPPs), which become apparent in trophozoite stages approximately 12-15 h post-invasion of the merozoite into the red blood cells (RBC) (Kirk 2001). The NPPs allow metabolic and biosynthetic substrate uptake of inorganic and organic ions, amino acids, sugars and nucleosides (Kirk 2001, Staines et al. 2004b, 2007, Bokhari et al. 2008); currently, parasite-induced permeability pathways are considered attractive chemotherapeutic targets (Staines et al. 2005, Kelly et al. 2007, Lopez & Segura 2008). Solanum nudum Dunal (Solanaceae) is a plant used in traditional medicine in Colombia, from which four cholest-4-ene-3,22-diones similar to progesterone steroids [tumacone A (SN1), tumacone B (SN2), tumacoside A (SN3), tumacoside B (SN4) and diosgenone] have been isolated and their structures elucidated (Sáez et al. 1998). These compounds have shown in vitro antiplasmodial activity against P. falciparum strain FCB-2 (Pabón et al. 2002). This report demonstrates that compounds from S. nudum are not mutagenic (Pabón et al. 2003), clastogenic (Alvarez et al. 2004) or cytotoxic to mammalian cells (Londono et al. 2006). Alterations of RBC shape and membrane properties are likely to interfere with the growth of parasites (Ziegler et al. 2004). One previous reported has shown that steroids may influence the biological membrane function of RBCs by modifying ion-permeability, metabolite transport, membrane microdomains, cell shape and energy metabolism (Haines 2001). In this study, we assessed the effect of steroids from S. nudum (SNs) on the membranes of noninfected red blood cells (nRBCs), the parasite invasiveness and the solute-induced lysis of pRBCs. We found that the majority of merozoites were unable to invade SN-treated erythrocytes; that SN2 and diosgenone did not affect the fine structure of nRBCs; and that none of the SNs were haemolytic. Additionally, SN2 and SN4 inhibited the solute-induced haemolysis of pRBCs. MATERIALS AND METHODS Collection of plants - Leaves of S. nudum, commonly named zapata, were previously collected in Tumaco (Nariño-Colombia) (78º30'W 1043'15"N, 15 m altitude) with the help of local healers and natives. S. nudum is a 2 m high shrub with white flowers, yellow stems and green fruit, which grows as a weed. Species confirmation was carried out at the Herbariums of Universidad Antioquia, Universidad Nacional and Universidad de Nariño. This was further confirmed by Professor Nee, New York Herbarium. Samples were prepared and deposited at the Herbarium of the Universidad de Antioquia (Colombia) under voucher 554 file 61181 of the Colombian flora catalogue. Compound isolation - The isolation and chemical structures of steroids SN1, SN2, SN4 and diosgenone have previously been reported (Sáez et al. 1998) (Table). In brief, ground, air-dried leaves were extracted by percolation with petroleum ether, then with MeOH and finally with AcOEt. The resultant extract was passed through a silica chromatography column with an elution system of CH2Cl2/AcOEt and evaluated by TLC. Structural verification of the steroids was made by spectroscopy and 1H-NMR. Test compounds were dissolved in dimethyl sulfoxide and diluted over different concentrations. P. falciparum culture - The chloroquine-resistant P. falciparum strain FCB-2/Colombia (Instituto Nacional de Salud, Bogotá) was grown in continuous culture by standard techniques at 37ºC with a gas mixture of 5% CO2, 5% O2 and 90% N2 (Trager & Jensen 1976). Parasite cultures were synchronised by 5% sorbitol treatment as described by Lambros and Vanderberg (1979) and a 60% Percoll gradient method was used to concentrate schizont-parasitised erythrocytes 36-40 h after invasion (Dluzewski et al. 1984). The percentage of pRBCs was determined by counting 1,000 erythrocytes in a thin blood smear stained with Giemsa and viewed under a light microscope (X100 objective). Effects of SNs on parasite invasiveness - Noninfected erythrocytes were treated with IC50 concentrations of SNs (Table) at 37ºC for 1 h. Excess SNs were thoroughly removed by washing RBCs twice in RPMI 1640 incomplete medium. Next, P. falciparum FCB-2 cultures rich in schizont containing RBCs (more 50%) were mixed with SN-pretreated erythrocytes. Cultures at the final haematocrit of 2% were then incubated at 37ºC for 20 h, at which time microscopical examination was carried out. Haemolysis assay of nRBCs - The haemolytic activity of SN compounds was assayed in fresh human type A+ RBCs that were washed three times in phosphate-buffered saline. RBCs were suspended at 5% haematocrit in an isosmotic HEPES-Tris buffer solution (10 mM HEPES-Tris, 5 mM glucose, 5 mM KCl, 1 mM EDTA, pH 7.4) containing 300 mM sorbitol (320 + 1 mmol Kg·H2O-1) or 150 mM NaCl (304.5 + 5.5 mmol Kg·H2O-1). The cellular suspension was incubated with SN and diosgenone steroids for 1 h and complete haemolysis was obtained using 1% Tween 20 (100% haemolysis). SN concentrations that were tested in this assay were equivalent to the IC50 reported for inhibition of parasite growth (Pabón et al. 2002) (Table). After incubation, samples were centrifuged (500 g, 5 min) and 100 μL of the supernatants were transferred to a flat-bottom 96-well microtitre plate. Released haemoglobin was monitored by measuring absorbance at 540 nm (Bio-Rad 610). The percentage of haemolysis was calculated as described previously (Efron et al. 2002). Effect of SNs on solute-induced haemolysis of P. falciparum-infected erythrocytes - The effect of SNs on parasite-induced permeability pathways was investigated by haemolytic assays in an isosmotic-solute solution as described previously (Go et al. 2004). Trophozoite-infected RBCs (more 50%) from a synchronised P. falciparum strain FCB-2 culture were washed twice with RPMI incomplete medium and resuspended in a solution of NaCl (150 mM) - HEPES (10 mM) (pH 7.4; 304.5 + 5.5 mmol Kg·H2O-1) at 50% haematocrit. Eleven microlitres of SN stock solution were combined with 88 μL of solution containing 300 mM solute (sorbitol or alanine) in 10 mM HEPES (pH 7.4; 320 + 1 mmol Kg·H2O-1) and 11 μL of pRBC suspension in NaCl-HEPES solution. After 30 min incubation at 37ºC, cell suspensions were centrifuged (500 g, 5 min) and supernatants (100 μL) were transferred to a flat-bottom 96-well microtitre plate. The released haemoglobin was read at 540 nm (Model 610 Microplate Reader, Bio-Rad) and the percentage of haemolysis was calculated as described previously (Go et al. 2004). Compounds associated with > 60% haemolysis inhibition were considered good inhibitors of the parasite-induced permeability pathways. Thus, the IC50 values (the concentration of compounds that reduced haemolysis to 50% of that measured in the absence of compound) were estimated by linear interpolation only for these compounds. The control was 100 μM Cl- channel blocker NPPB [5-nitro-2-(3-phenylpropylamino) benzoic acid], a well-known inhibitor of pRBCs NPPs (Huber et al. 2002). Analysis of nRBCs by transmission electron microscopy (TEM) - nRBCs were exposed to a concentration twice the IC50 value of each agent (251 μM of SN2, 43.6 μM of diosgenone and 0.15% DMSO as control) in RPMI incomplete medium and incubated at 37ºC for 1 h. Excess SNs were removed by washing RBCs twice with RPMI incomplete medium. RBCs were fixed for 1 h with 2.5% glutaraldehyde in 0.1 M sodium cacodylate buffer, pH 7.2. Cells were post-fixed with 1% osmium tetroxide in 0.1 M cacodylate buffer and dehydrated with acetone solutions. Samples were embedded in PolyBed (Polyscience Inc.) and cut into 50-60 nm-thick sections, stained (uranyl acetate and lead citrate) and examined by Zeiss 900 TEM. Data analysis and statistics - Data were analysed and plotted using GraphPad Prism version 4.03 for Windows (GraphPad Software, San Diego, California, USA). The one-way analysis of variance (ANOVA) test was used for statistical analysis followed by post hoc comparison using Dunnett's Multiple Comparison Test. P values < 0.05 were considered significant. At least two independent experiments were performed for each assay. RESULTS Effect of SNs on parasite invasion of RBC - To assess the effects of SNs on erythrocytes, SN-treated RBCs and schizont-infected RBCs were mixed and maintained in culture for 20 h. It was found that the schizonts were capable of rupturing erythrocytes while the merozoite invasion was significantly inhibited in treated RBCs (p < 0.05). The percentages of reduction of ring-infected RBCs were: 51% SN1, 50% SN2, 44.3% SN4 and 60% diosgenone (Fig. 1). SNs and diosgenone are non toxic to nRBCs - Due to the above observations, we examined the haemolytic activity of the SNs against nRBCs as a measure of toxicity. The data showed that neither the SNs nor 0.15% DMSO caused significant isosmotic haemolysis at the concentration evaluated. The observed hemolysis was less than 1%. To determine if there is a relationship between the invasion inhibition and alterations of RBC morphology, TEM assays of nRBCs treated with 251 μM SN2 or 43.6 μM diosgenone were performed. No obvious effect of SN2 and diosgenone was observed on the morphology of nRBCs (Fig. 2). Inhibition of the solute-induced lysis of pRBCs by SN2 and SN4 - The compounds SN1, SN2, SN4 and diosgenone were assayed for their ability to inhibit the parasite-induced permeability pathways. The concentrations used in these assays were their respective IC50 values for inhibition of parasite growth that have been previously reported (Pabón et al. 2002) (Table). Both diosgenone (21.8 μM) and SN1 (21 μM) were only able to inhibit solute-induced lysis by less than 20%, which was significantly less than 100 μM NPPB (p < 0.05) (Fig. 3). In contrast, 125.5 μM SN2 and 57.4 μM SN4 inhibited the sorbitol and alanine-induced lysis by more than 80% and 50%, respectively, relative to the control without SNs. Thus, IC50 + SEM were estimated for compounds SN2 and SN4, which inhibited solute-induced lysis in a dose-responsive manner with similar potency (p > 0.05) (Table). These concentrations were similar for both solutes and lower than those obtained for P. falciparum in vitro growth inhibition (Table). The inhibition with 100 μM NPPB was 89.8% and 68.4% in isosmotic sorbitol and alanine, respectively (Fig. 3). Haemolysis of pRBCs in isosmotic 150 mM NaCl was completely sensitive to NPPB, but neither the SNs nor diosgenone inhibited the NaCl-induced lysis. DISCUSSION Steroidal compounds have a broad spectrum of biological effects and, because of their chemical structure, they can exert effects on the lipid bilayer of cells or at the intracellular level (Haines 2001); therefore, these molecules have pharmaceutical potential. It has been previously demonstrated that the antiplasmodial activity of compounds such as lupeol (Ziegler et al. 2002), betulinic acid and its analogues (Ziegler et al. 2004), triterpenoid saponins from Glinus oppositifolius (Traore et al. 2000) and dehydroepiandrosterone (DHEA) and its analogues, 16alpha-bromoepiandrosterone (Freilich et al. 2000), may be explained by their action on the membranes of RBCs. It has been demonstrated that amphiphilic and membrane-modifying agents can inhibit P. falciparum growth, with good correlation to IC50, by inducing changes in membrane curvature, which results in stomatocytes or echinocytes. It has been suggested that this is due to their incorporation into erythrocyte membranes (Ziegler et al. 2004, Ziegler et al. 2002). We found a significant reduction in the percentage of ring-infected RBCs after mixing schizonts with SN-treated erythrocytes, suggesting that SNs may partially inhibit merozoite invasion of erythrocytes. Although diosgenone and SN2 did not change the shape of uninfected RBCs, an effect on cell deformability and/or rigidity cannot be excluded because TEM only provides fine cell structure; therefore, other studies are necessary to more closely examine the effects of SNs on RBCs. Moreover, data showed that the SNs are not haemolytic to normal RBCs and the cytotoxic activity of SNs on RBCs can therefore be discounted. The effect of the SNs on parasite invasion may be due to their interaction with the erythrocyte surface or erythrocyte membrane and studies of differential scanning calorimetry using the SNs in artificial membranes are currently ongoing. After invasion by P. falciparum, the erythrocyte membrane undergoes functional modifications, which include changes in RBC membrane lipid composition and exposure of parasite proteins on the surface of the cell host plasma membrane (Kirk 2001, Marchesini et al. 2005). During its development inside the RBC, the parasite increases the host cell membrane permeability to allow for uptake of nutrients, elimination of waste products, volume regulation and control of ion concentration within the erythrocytes (Staines et al. 2005). One method for determination of the altered pRBC permeability involves suspending cells in an isosmotic solution, sorbitol for example, because the RBC membrane is normally impermeant to this solute (Go et al. 2004, Kirk 2001). This method provides accurate and informative data regarding pRBC permeability (Staines et al. 2005). Although the nature of the NPP is debated, it is considered a chemotherapeutic antimalarial target (Lopez & Segura 2008, Staines et al. 2005) and the relationship between inhibition of permeability pathways and growth inhibition of different substances has been reported (Go et al. 2004, Kang et al. 2005, Lisk et al. 2006, Staines et al. 2004a), although to date they have not been fully correlated (Go et al. 2004, Huber et al. 2002, Kang et al. 2005). We found that SN2 and SN4 need IC50 values less than that needed for inhibition of parasite growth to inhibit the permeability pathways of pRBCs. This suggests that the antiplasmodial activity of SN2 and SN4 may be partially related to the inhibition of the NPP on the pRBC and therefore may be their mechanism of action. Inhibition of solute-induced haemolysis was comparable for SN2, SN4 and NPPB in both sorbitol and alanine. In contrast, in isotonic NaCl, SN2 and SN4 failed to inhibit haemolysis whereas NPPB was effective. Moreover, the haemolysis in pRBCs was less in isosmotic sorbitol with NPPB as compared with NaCl without NPPB, suggesting a significant uptake of NaCl and haemolysis in pRBCs exposed to isotonic saline, which is similar to a previous report (Huber et al. 2002). This differential effect can be explained because NPPB is a Cl- channel blocker and not an exclusive inhibitor for the NPP, while SN2 and SN4 could potentially be acting more specifically on NPPs. The inhibition of solute-induced haemolysis of pRBCs in alanine and sorbitol was similar for SN2 and SN4. This data is in agreement with a report from Kirk et al. (1994), who demonstrated that the time course for haemolysis of pRBCs in isosmotic alanine and sorbitol was comparable, suggesting a similar mechanism of induced transport in P. falciparum-infected pRBCs (Kirk 2001, Kirk et al. 1994, Staines et al. 2004b). Although solute-induced haemolysis does not reflect in vivo conditions of pRBCs, the observed inhibition by SN2 and SN4 suggests effects of SNs on pRBC permeability, which could interfere with parasite development by closing the routes used to dispose of waste and take up nutrients. In contrast, the antiplasmodial activity of SN1 and diosgenone differs from that observed for SN2 and SN4 and this mechanism is currently under study. ACKNOWLEDGMENTS To Lina Ortiz and Ana María Mesa, for their technical assistance in SN preparation, to Dr. Rossiane Vommaro, from the Laboratório de Ultraestrutura Celular Hertha Meyer, Universidade Federal do Rio de Janeiro, Brazil, for her assistance with TEM work, to Mariano Zalis, Laboratório de Infectologia e Parasitologia Molecular, Universidade Federal do Rio de Janeiro, and Dr. Henry Staines, for his valuable critical reading of the first draft and suggestions for complementary assays. REFERENCES
Copyright 2009 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc09154t1.jpg] [oc09154f2.jpg] [oc09154f1.jpg] [oc09154f3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}