 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
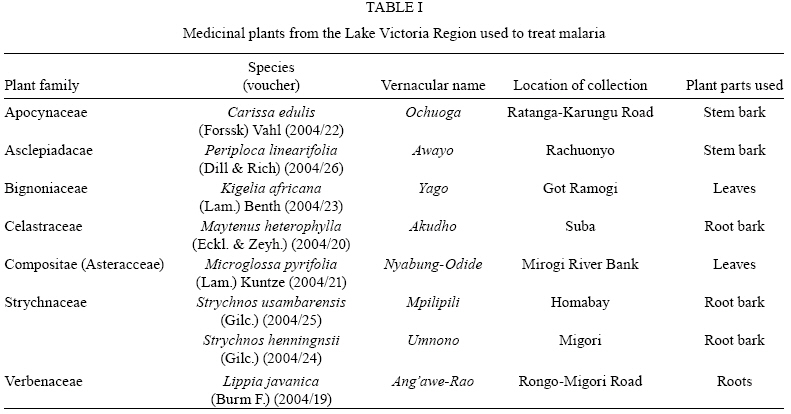
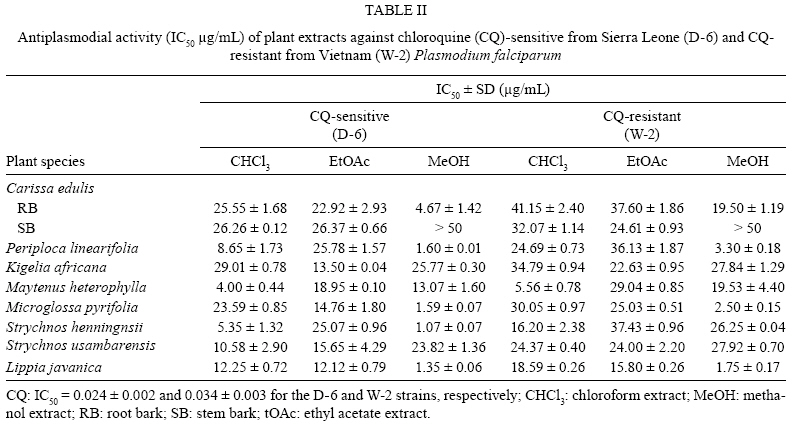
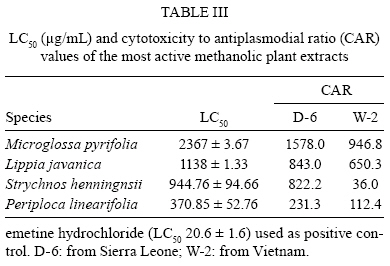
Memórias do Instituto Oswaldo Cruz, Vol. 104, No. 5, August, 2009, pp. 689-694 ARTICLES In vitro antiplasmodial activity and toxicity assessment of plant extracts used in traditional malaria therapy in the Lake Victoria Region Teresa Akeng'a AyukoI, +; Richard Njihia NjauII; Wanjala CorneliusI; Nyangasi LeahII; Isiah Omollo NdiegeIII IDepartment
of Applied Chemistry, Kigali Institute of Science and Technology, BP 3900 Kigali,
Rwanda Financial support: SIDA, VICRES, IUCEA Received 2 December
2008 Code Number: oc09155 ABSTRACT As part of our program screening the flora of the Lake Victoria Region, a total of 54 organic extracts from seven plant families (8 species) were individually tested for antiplasmodial activity against chloroquine-sensitive [Sierra Leone (D-6)] and chloroquine-resistant [Vietnam (W-2)] strains. Only 22% of these extracts exhibited very high in vitro antiplasmodial activity. Six methanol (MeOH) extracts and one chloroform extract showed in vitro antiplasmodial activity against the D-6 Plasmodium falciparum strain, while only three MeOH extracts were active against the W-2 strain. All of the ethyl acetate extracts proved to be inactive against both strains of P. falciparum. A brine shrimp cytotoxicity assay was used to predict the potential toxicity of the extracts. The cytotoxicity to antiplasmodial ratios for the MeOH extracts were found to be greater than 100, which could indicate that the extracts are of low toxicity. Key words: antiplasmodial - cytotoxicity - plant extracts - malaria therapy Eighty percent of the world's population uses herbal remedies (Zirihi et al. 2005). There are several plants that have been used by people living around the Lake Victoria Basin, a malaria endemic zone, to manage and treat malaria (Gachathi 1989, Kokwaro 1993). There is a need to carry out systematic investigations using modern scientific methods to establish the efficacy and toxicity of the medicinal plants commonly used by traditional medical practitioners. Thus, organic extracts from eight plants used by local indigenous communities in the Lake Victoria Region for traditional malaria therapy, namely Microglossa pyrifolia (Lam.) Kuntze, Kigelia africana (Lam.) Benth, Lippia javanica (Burm F.) Spreng, Carissa edulis (Forssk) Vahl., Maytenus heterophylla, (Eckl. & Zeyh.)Strychnos henningnsii (Gilc.), Strychnos usambarensis (Gilc.) and Periploca linearifolia (Dill and Rich), were studied in regard to their antiplasmodial activity and were evaluated for toxicity against brine shrimp larvae (Artemia salina). MATERIALS AND METHODS Plant collection - The plant materials (Table I) were collected from the Lake Victoria Basin in Nyanza Province, Kenya, in May 2003. Their selection was based on documented literature and interviews with local communities (Kokwaro 1993). The plant specimens were identified by S.G. Mathenge of the University of Nairobi Herbarium, where voucher specimens were deposited. The samples were cut into small pieces and air-dried in the shade for two weeksand then ground into a fine powder using a motorised laboratory grinding mill (Christy and Norris Ltd, Chelmsford, England). Extraction - All of the solvents used were of analytical grade. For each sample of ground plant material (100 g), sequential cold extractions were performed for 72 h using hexane, chloroform (CHCl3), ethyl acetate (EtOAc) and methanol (MeOH). The extracts were decanted and filtered through Whatman filter paperand the resulting filtrates were dried under reduced pressure (40ºC). The percentage yield for each sample was determined and the crude extracts were stored in a freezer at -4ºC. Preparation of drugs - Stock solutions of the crude extracts (10 mL, 250 μg/mL) were made with sterile de-ionised water and filtered through 0.45 and 0.22 μm micro-filters in a laminar flow hood. The water insoluble extracts were first dissolved in dimethylsulphoxide (DMSO) (solvent concentration < 0.025%) (Elueze et al. 1996) and then sonicated until a homogeneous solution was obtained. Stock solutions of the reference drug [chloroquine (CQ) sulphate salt, purity > 99%, Sigma-Aldrich Kenya Ltd] were similarly prepared in sterile water. All of the drugs were stored under refrigeration. Cultures of Plasmodium falciparum - Two laboratory adapted P. falciparum cultures, D-6 (CQ-sensitive from Sierra Leone) and W-2 (CQ-resistant from Vietnam) were used in the study. The strains were cultured and maintained at the Walter Reed Malaria Research Laboratories at the Kenya Medical Research Institute (KEMRI), Nairobi. The culture medium was a variation of that described by Trager and Jensen (1976), which consisted of RPMI 12640 supplemented with 10% human serum, 25 mM N-2- hydroxyethylpiperazine-N-2-ethanesulfonic acid and 25 mM NaHCO3. Human type O+ erythrocytes served as host cells. The cultures were incubated at 37ºC in an atmosphere of 3% CO2, 5% O2 and 92% N2. Antiplasmodial assay - The antiplasmodial assays were carried out at the Walter Reed Army Institute of Research laboratories based at KEMRI. An in vitro semi-automated micro-dilution assay technique was used that measures the ability of the extracts to inhibit the incorporation of [G-3H] hypoxanthine into the malaria parasite (Desjardins et al. 1979, O'Neill et al. 1985). Aliquots of the culture medium (25 μL) were added to all the wells of a 96-well flat-bottomed micro-culture plate (Costar Glass Works, Cambridge, UK). The test solutions (25 μL) were added in duplicate to the first wellsand a Titertek motorised hand diluter (Flow Laboratories, Uxbridge, UK) was used to make two-fold serial dilutions of each sample over a 64-fold concentration range. The susceptibility tests were carried out with an initial 200 μL of parasite culture (0.4% parasitaemia, 1.5% haematocrit) in each well. The stock solutions of the plant extracts were diluted in the plate to give a 100 μg/mL concentration (as the highest concentration) and then diluted two-fold until reaching a concentration of 1.56 μg/mL. A suspension (200 μL, 1.5% v/v) of parasitised erythrocytes (0.4% parasitaemia) in the culture medium was added to all the test wells. Non-parasitised erythrocytes were used in control experiments. The plates were incubated at 37ºC in an atmosphere of 3% CO2, 5% O2 and 92% N2. After 48 h each well was pulsed with 25 μL of culture medium containing 0.5 μCi of [G-3H]-hypoxanthineand the plates were incubated for a further 18 h. The contents of each plate were harvested onto glass fibre filters, washed thoroughly with distilled water, dried and the radioactivity measured using a BetaplateTM liquid scintillation counter (Wallac, Zurich, Switzerland). Data from the counter was imported into a Microsoft Excel spreadsheet, which was then transferred into an Oracle database programme to determine IC50 values. A minimum of three separate determinations was carried out for each sample. Brine shrimp toxicity (BST) assay - BST assays were carried out on the MeOH extracts that showed good antiplasmodial activity. The brine shrimp (A. salina) toxicity bioassay was conducted according to McLaughlin et al. (1991). Artificial seawater was prepared by dissolving sea salt (38.0 g) in 1 L of distilled water. The seawater was put in a small tank and a teaspoon of brine shrimp eggs was added to the covered half of a divided tank. The other side was not covered so as to allow light in that attracted the hatched shrimps. The tank containing the brine shrimp eggs was left at RT for 48 h to allow the eggs to hatch. Samples of the extracts were dissolved in DMSO and diluted with artificial seawater so that the final concentration of DMSO did not exceed 0.05%. Different concentrations (2000, 1000, 500, 400, 200, 100, 50 μg/mL) of the MeOH extracts were prepared, in triplicate, in test tubes. The test tubes used were washed and sterilised in an autoclave. Brine shrimp larvae (nauplii, 10) were added to each tube. All the test tubes were covered at RT for 24 h. After this period, the number of dead and surviving brine shrimps was recorded. Each experiment was replicated three timesand the average values were determined. The toxicity of each extract was determined from 50% lethal concentration values determined by probit analysis (Solis et al. 1993). Emetine chloride (98% purity, Sigma-Aldrich Kenya Ltd) was used as a positive control. RESULTS Table I lists the plants that were identified as being used in the treatment and management of malaria-related fevers. The table includes the plants' family, species, voucher number, parts used, local names and the exact location of the sampling. The results of the in vitro antiplasmodial tests are summarised in Table II. Two strains of Plasmodium were used, a CQ-sensitive strain and a CQ-resistant strain. CQ was used as a positive control. The activities of the crude extracts were grouped according to Deharo et al. (2001). Thus, any extracts that showed an IC50 for antiplasmodial activity of less than 5 μgmL-1 were considered active, while those that had IC50's from 5-10 μgmL-1 were considered moderately activeand those extracts with IC50's of over 10 μgmL-1 were considered inactive. The activities obtained indicate that there are differences in the strains' sensitivities to the extracts. The hydrophilic MeOH extracts of P. linearifolia, M. pyrifolia and L. javanica showed high anti-parasitic activity against both strains of P. falciparum. However, the methanolic extract from S. henningnsii was active against the CQ-sensitive strain and inactive against the CQ-resistant strain of P. falciparum. The CHCl3 extracts from M. heterophylla had moderate activity towards both strains, but the CHCl3 extracts from S. henningnsii were only moderately active against the CQ-sensitive P. falciparum strain. All of the EtOAc extracts were inactive, as well as the extracts from the stem bark (SB) of C. edulis. The methanolic extracts from this specie's root bark (RB) were moderately active against the D-6 strain of P. falciparum, but were inactive against the W-2 strain. It has been reported elsewhere that the methanolic extracts of C. edulis and S. henningnsii roots obtained from Central Kenya (Meru) have mild antiplasmodial activity against CQ-sensitive (ENT 30) and CQ-resistant (NF 50) strains of falciparum (Kirira et al. 2006). Well-known traditional antimalarial plants like K. africana and S. usambarensis were found to be inactive in vitro. This phenomenon can be explained by several factors, such as the degradation of active compounds during storage or the presence of prodrugs that undergo enzymatic transformations in vivo to give active antiplasmodial compounds (Krettli et al. 2001). During this study, it was not feasible to subject all the lipophilic hexane extracts to antiplasmodial assays. This was mainly due to low yields and insufficient quantities of reagents. The only hexane extract assayed was from L. javanica and that extract was found to be inactive. The brine shrimp toxicity data and the calculated cytotoxicity to antiplasmodial ratios (CARs) are summarised in Table III. The methanolic extracts from the four plant species with the highest antiplasmodial activities were subjected to the BST assay. The methanolic extracts from these antiparasitic plants exhibited no significant toxicity. The M. pyrifolia extract was found to be the least toxic. Emetine hydrochloride, a standard antiparasitic therapy, was found to be more toxic to the shrimps than any of the methanolic plant extracts tested. However, further tests, like in vivo cytotoxicity measurements or in vitro tests with HeLa cells are needed to conclusively determine these extracts' CAR values. DISCUSSION During rational drug discovery from plant sources, in vitro and/or in vivo assays are carried out. In vitro assays are performed in order to determine the activity of the target molecules before toxicity studies in the real organism are performed. While in vivo tests are generally more practical, more rapid and less expensive than in vitro cultures, not all anti-malarial drugs are active in the Plasmodium berghei mouse model (Ferreira-da-Cruz et al. 2000). Also, in vivo studies require significantly larger quantities of the potential therapeutic agent when compared to in vitro assays, such that about 1 g of extract is required per in vivo test (Krettli et al. 2001). The selection of plant species for the study (Table I) was based on ethnopharmacology and documented literature. Although a random selection of plants might have been used, this procedure was considered inadequate. Indeed, it has been reported that by employing a random selection, only 1% of plants among 300 randomly selected species were found to be active, while nearly 20% of about 50 plants selected in light of traditional knowledge were found to be active (Krettli et al. 2001). There was a large variability in the activity of the crude extracts against P. falciparum (Table II). The distribution of active ingredients in any plant is uniqueand depends on the species, age, geographical variations and the specific plant tissue used. Traditional practitioners use different plant parts from each species according to their efficacy. However, the plant extracts investigated during this study can be largely classified into two groups. The first group includes all the extracts that showed a high activity (IC50 < 10μg/mL, Table II). This group of extracts (22%) may be useful in the treatment of malaria and thus further purification and isolation of the active compounds from this group of extracts and their development into drugs should be explored. The second group of extracts has IC50's > 20 μg/mL (56%). Additionally, there are a few borderline extracts with IC50 values > 10 μg/mL. The activity values obtained in this study are comparable to those of known plant remedies of various origins (O' Neill 1987, MacKinnon et al. 1997, Kristina et al. 1999, Oketch-Rabah et al. 1999, Krettli et al. 2001, Bertani et al. 2005, Vigneron et al. 2005, Zirihi et al. 2005, Chikara et al. 2006, Kirira et al. 2006, Mesquita et al. 2007). For example, when studying the crude ethanolic extracts of Artemisia annua by our study's protocol, the IC50 value against the CQ-resistant K-1 strain was found to be 3.9 μgmL-1 (O' Neill et al. 1985). The observed activities of the hydrophilic extracts are in contrast to those reported by Jenett-Siems et al. (1999), who found that lipophilic extracts were more active than hydrophilic extracts. Hydrophilic extracts are closer in composition to the aqueous preparations commonly used by traditional practitioners. François et al. (1996) have similarly investigated the organic extracts from the roots, SB, fruit rind, seeds and leaves of plantsand found a wide range of activities with the highest activities being found in the methylene chloride root extracts. Several classes of plant secondary metabolite are responsible for the observed anti-malarial activity, but the most important and diverse bio-potencies have been observed in alkaloids, quassinoids and sesquiterpene lactones (Quetin-Leclercq et al. 1991, Wright et al. 1994, Schwikkard & van Heerden 2002, Saxena 2003). The plants investigated during this study belong to various families, from which similar metabolites have been isolated. For instance Andrade-Neto et al. (2007) isolated antiplasmodial indole alkaloids from plant species of the Apocynaceae family in the Brazilian Amazon. Several phytochemical reports have been published concerning extracts from the four plants we found to be most active. The reports on these medicinal plants indicate that the plant species contain antiplasmodial metabolites. The antiplasmodial alkaloids isostrychnofoline (IC50 1.2 μM) and holstiine (IC50 1.0 μM) have been isolated from the RB of S. henningnsii Strychnaceae (Frederich et al. 1999, 2002). Most plants from the Celastraceae family contain a class of sesquiterpene nicotinoyl alkaloids, which are characterised by a dihydroagarofuran sesquiterpene bonded as a diester to a substituted nicotinic acid (Schaneberg et al. 2001). 1β-Acetoxy-9α-benzoyloxy-2β, 6α-dinicotinoyloxy-β-dihydroagarofuran and β-amyrin have been isolated from M. heterophylla (Orabi et al. 2001). The leaf extracts from P. linearifolia, in the family Asclepiadacae, contain phytin, anthraquinone, gemnamagenin, gymnemic acid II, betaine, cholin and lupeol (Nagaraju & Rao 1990, Hichri et al. 2002). Additionally, M. pyrifolia, Compositae (Asteraceae), leaves have been reported to contain several terpenoids, such as E-phytol, 1,3- hydroxyoctadeca-9Z, 11E, 15-trien oic acid and 6E-geranylgeraniol-19 oic acid, which exhibit antiplasmodial IC50 values between 2.5-13.7 μg/mL (Köhler et al. 2002). The reported data on L. javanica (Verbenaceae) indicates that there are several terpenoids isolated from the flowering tops of this species. These terpenoids exhibit larvicidal, antimalarial, spasmolitic, sedative, hypotensive and anti-inflammatory activities (Pascual et al. 2001, Omolo et al. 2004). However, there is no phytochemical data reported on root extracts from L. javanica. The CAR was very high for most of the extracts tested. These CAR values are comparable to those reported for CQ (518.27) on HeLa cells after 72 h (Yapi et al. 2000). This may indicate the selectivity of these extracts because the extracts exhibit weak toxicity and good CAR values (Munoz et al. 1999). However, several reports demonstrate that the antimalarial activity of a compound and its toxicity are not directly related (O'Neill et al. 1987, Schwikkard & van Heerden 2002, Kirira et al. 2006). There are reports that certain other plant species' methanolic extracts exhibit significant toxicity in the brine shrimp assay (Mohtasheen et al. 2001, Kirira et al. 2006). In conclusion, only 22% of the 54 organic extracts from plants used in traditional malaria therapies in the Lake Victoria Region showed high antiplasmodial activity. Methanolic extracts from several species were found to exhibit high antiplasmodial activityand low toxicity toward test organisms. In this study a shrimp assay was used for toxicity evaluation, we recommend that further toxicity tests be conducted on HeLa cell lines under conditions similar to the antiplasmodial test in order to better determine these extracts CAR values. ACKNOWLEDGEMENTS To the Kenya Medical Research Institute (KEMRI) and US Army Walter Reed Research Institute, for allowing us to use their laboratory equipment, and to the technical support from Liyala Pamela and Akala Hosea. REFERENCES
Copyright 2009 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc09155t1.jpg] [oc09155t2.jpg] [oc09155t3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}