 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 104, No. 5, August, 2009, pp. 728-735 ARTICLES Simulium (Inaequalium) marins, a new species of black fly (Diptera: Simuliidae) from inselbergs in Brazil Mateus PepinelliI, +; Neusa HamadaII; Douglas C CurrieIII ILaboratório
de Entomologia Aquática, Faculdade de Filosofia, Ciências e Letras
de Ribeirão Preto, Universidade de São Paulo, Ribeirão
Preto, SP, Brasil Financial support: BIOTA/FAPESP (2007/56574-4), MCT/CNPq (NH) Received 13 January
2009 Code Number: oc09163 ABSTRACT A new species of black fly, Simulium (Inaequalium) marins (Diptera: Simuliidae), is described based on the male, female, pupa and larva. This new species was collected from two localities: a small stream on the Pico dos Marins, a high mountain with granite outcrops in Piquete County, state of São Paulo, and in a small stream in the Serra dos Órgãos National Park, state of Rio de Janeiro, Brazil. Key words: aquatic insects - Simulium margaritatum - Simulium rappae - Pico dos Marins - São Paulo The Neotropical subgenus Simulium (Inaequalium) Coscarón & Wygodzinsky includes 15 nominal species, of which 13 occur in Brazil (Adler & Crosskey 2009). Species in the subgenus S. (Inaequalium) are typically associated with pristine mountain streams and have been recorded at a wide range of altitudes (Pepinelli et al. 2005b, 2006). The two most recently described species were collected from high-altitude streams associated with inselbergs in Chapada Diamantina National Park (Pepinelli et al. 2006) and Pico da Neblina National Park (Py-Daniel & Barbosa 2007). Inselbergs are isolated rock outcrops that rise abruptly from the surrounding terrain. Granitic inselbergs are geologically and geomorphologically old, representing remnants left by erosion processes; they range in elevation from a few to several hundred meters (Coor et al. 1993). Occurring over a broad spectrum of climatic zones (Porembski 2007), inselbergs have received considerable attention in recent years because they represent islands of high biotic diversity (Burke 2001). Such areas support a variety of microhabitats, each with its own distinct community (Porembski et al. 2000). Many aquatic and semi-aquatic invertebrates are confined to permanent or temporary watercourses that issue from these rock outcrops. In this paper we describe a new species of black fly in the subgenus S. (Inaequalium) based on specimens collected from two small inselberg streams located in Southeastern Brazil. MATERIAL AND METHODS All material used in this description was collected in one stream, located on Marins Peak (Pico dos Marins), a 2,200 meter high granitic outcrop (Figs 1, 2) situated in Piquete County, state of São Paulo (SP), Brazil, near the border with state Minas Gerais (MG). This peak is part of the Serra da Mantiqueira mountain range of Southeastern Brazil. We also collected pupal exuviae from a granitic outcrop located in the Serra dos Órgãos National Park, in a small stream at an altitude of 1,580 m in Teresópolis County, state of Rio de Janeiro (RJ), which is part of the Serra do Mar mountain range of Southeastern Brazil. Collection and rearing techniques are described in detail by Pepinelli et al. (2005a). Measurements and terminology for species description follow Adler et al. (2004) and Hamada and Pepinelli (2004). Description of the adult thoracic pattern was based on specimens recovered from alcohol using the technique of Sabrosky (1966). Morphological features were photographed using a Leica digital camera attached to a dissecting or to a compound microscope. Type specimens are deposited in the Zoology Museum, at São Paulo University (MZUSP), SP, Brasil. Other paratypes are deposited in the Invertebrate Collection of the National Institute of Amazonian Research (INPA), Manaus, Amazonas, Brazil. Simulium
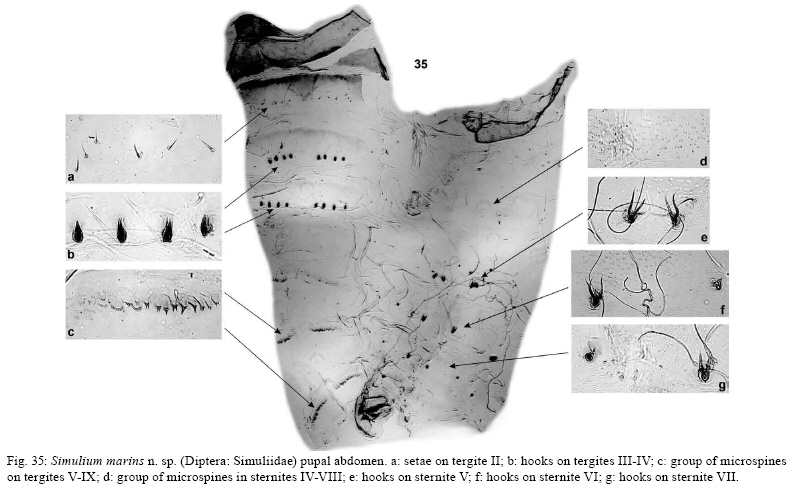
(Inaequalium) marins Female: (reared from pupa, n = 1) (Figs 3, 4, 5-16). General body color dark brown to black (Figs 3, 4). Body length 2.4 mm; thorax lateral length 0.96 mm; thorax dorsal length 0.91 mm. Wing length 2.6 mm; wing width 1.2 mm. Fronto-ocular triangle well developed (Fig. 6). Frons dark gray, clypeus and occiput light gray, with silver pruinosity. Antenna with silver pubescence, 0.52 mm in length; scape, pedicel and base of first segment light brown, remaining segments dark brown (Fig. 5). Maxillary palpus brown; sensory vesicle elongated occupying about 1/2 of the length of palpomere III; palpomere V twice the length of palpomeres III and IV (Fig. 13). Mandible with six external serrations and 30 internal teeth. Lacinia with 26 retorse teeth. Cibarium with well-developed, sclerotized cornuae, with teeth arranged over median elevation (Fig. 8). Thorax with scutum black covered by evenly arranged, recumbent, golden greenish hairs, without pattern (Figs 3, 4). Anepisternum and katepisternum gray with silver pruinosity. Scutellum light brown with long dark setae and golden hairs. Postnotum with golden pruinosity. Costa of wing with spines and setae. Subcosta with sparse distribution of setae. Basal section of Radius bare; R1 with spines and setae; Rs with setae. Fore leg with coxa, trochanter and femur light brown, tibia with apical and basal region brown and middle region whitish, basitarsus and tarsomeres I-IV dark brown (Fig. 10). Middle leg with coxa, trochanter and femur brown, proximal 1/3 of tibia whitish and distal 2/3 dark brown, more than half of the proximal region of basitarsus whitish, remaining dark brown and tarsomeres I-IV dark brown (Fig. 11). Hind leg with coxa and trochanter brown, femur whitish with distal region dark brown, proximal 3/4 of tibia whitish and distal 1/4 dark brown, proximal 3/4 of basitarsus whitish and distal 1/4 dark brown, tarsomeres I-IV dark brown (Fig. 12). Calcipala reaching pedisulcus. Femur and tibia of legs with scale-like setae distributed among the filiform ones. Claws curved with sub-basal tooth (Fig. 7). Abdominal tergite II with silver pruinosity on anterolateral margins (Fig. 3). Tergites plates II-IX with varnish-like appearance. Basal fringe with long, golden hairs. Sternites whitish brown; genitalia dark brown. Eighth sternite weakly sclerotized with evenly distributed setae; hypogynial valves (= gonapophyses) sub-triangular, membranous and with few microtrichia, except weakly sclerotized on internal margins (Figs 9, 16). Cercus rounded, covered with small hairs and long dark setae; anal lobe (= paraproct) sub-triangular, with a small protuberance near the cercus (Figs 9, 15). Genital fork stout and sclerotized; lateral arms well developed, with a large anterior process; lateral arms forming sub-oval space (Fig. 14). Spermatheca globular, with internal spicules arranged in groups; spermathecal duct unpigmented. Male: (reared from pupa, n = 1) (Figs 17-18, 19-26). General body color dark brown (Figs 17, 18). Body length 2.9 mm; thorax lateral length 0.96 mm; thorax dorsal length 0.94 mm. Wing length 2.8 mm; wing width 1.3 mm. Antenna (Fig. 21) brown with silver pubescence, 0.45 mm in length. Palpus dark brown, sensory vesicle occupying less than 1/3 of palpomere III; palpomere V about 1.5 times as long as palpomeres III and IV (Fig. 22). Scutum black covered with golden hairs, without pattern (Figs 17, 18). Scutellum dark brown with long, black hairs and short, sparse golden hairs. Postnotum with silver pruinosity. Anepisternum and katepisternum gray with silver pruinosity. Costa of wing with spines and setae (Fig. 19). Subcosta with few setae (Fig. 19). Basal section of radius bare (Fig. 19); R1 with spines and setae; Rs with setae. Coloration of legs as the same in the female. Claws with well-developed thumb-like lobe. Calcipala reaching pedisulcus. Femur and tibia of middle and hind leg with scale-like setae distributed among the filiform ones. Abdominal tergites (Fig. 17) dark brown; basal fringe with long, thin, black hairs; tergite II black with two spots of silver pruinosity that extend to the side of the abdomen; tergite III without silver pruinosity; tergites IV-VII with dorsolateral silver pruinosity. Abdominal sternites membranous. Sternites III-VIII with silver pruinosity on median region. Genitalia dark brown; gonocoxite and gonostylus covered with small hairs and long setae; gonocoxite sub-quadrangular (Fig. 26); gonostylus subcylindrical (Figs 25, 26), elongated, narrowing apically, with one sub-terminal hook. Ventral plate covered by long hairs as in Fig. 23. Median sclerite Y-shaped. Paramere with developed and sclerotized basal process and numerous stout teeth (Fig. 24). Pupa: (n = 1) (Figs 27-34, 35). Cocoon length dorsally 3.3 mm, ventrally 3.3 mm; pupa lateral length 3.5 mm. Cocoon slipper-shaped, light brown composed of thick coalesced fibers with long dorsal projection (Figs 27-30). Cephalic plate covered by spines (Figs 30-32) with 2 + 2 long, bifid frontal and 1 + 1 bifid dorsal trichomes; rounded tubercles, distributed between the antennal shelters (Figs 31, 32). Thorax with spines on the anterior region (Fig. 33), five pairs of long, bifid trichomes near margin of dorsal cleft; tubercles rounded, sparsely distributed over surface. Gill length 2.1 mm (n = 2) pale brownish. Gill with six filaments (Fig. 34), with main trunk short, giving rise to three short sets of primary branches, each consisting of two secondary branches. All gill filaments approximately same length, with rounded ends. Abdominal tergites as in Fig. 35a-c. Tergite I with 2 + 2 filiform setae and 1 + 1 submedian short and strong setae. Tergite II with 6 + 6 filiform setae (Fig. 35a). Tergites III-IV with 4 + 4 submedian simple hooks (Fig. 35b) and 2 + 2 filiform setae. Tergite V with 3 + 3 small filiform setae. Tergites VII-IX with spine combs on anterior margin (Fig. 35c). Tergite IX weakly sclerotized. Abdominal sternites as in Fig. 35d-g. Sternites III-IV with spine combs on anterior margin (Fig. 35d) and 3 + 3 filiform setae. Sternite V with 2 + 2 close, bifid hooks in a row and 2 + 2 simple trichomes (Fig. 35e). Sternite VI with 1 + 1 bifid or trifid hooks, 1 + 1 small bifid hooks and 3 + 3 filiform setae (Fig. 35f). Sternite VII with 1 + 1 bifid or trifid hooks, 1 + 1 bifid hooks and 2 + 2 filiform setae (Fig. 35g). Sternite VIII without hooks. Sternite IX well sclerotized. Abdominal sternites III-VIII with spine combs. Larva: (last instar, n = 5) (Figs 36-44). Body length: 5.5-5.7 mm (mean = 5.6 mm, SD = 0.1); head capsule lateral length: 0.71-0.78 mm (mean = 0.76 mm, SD = 0.04); dorsal width of head capsule: 0.60-0.65 mm (mean = 0.63 mm, SD = 0.02). General body color light gray (in Carnoy's Fixative) (Figs 40, 41). Head pale to yellowish brown with positive pattern as in Fig. 36. Cuticle of dorsal and ventral region with small, simple setae. Cervical sclerites small, free in the membrane. Postgenal cleft narrow, semi-rounded (Fig. 37). Postgenal bridge as long as hypostoma (Fig. 38). Hypostoma with strongly pigmented anterior margin and nine apical teeth (Fig. 38). Median tooth simple and more prominent than sublateral teeth, 3 + 3 rows of sublateral teeth, the middle tooth smaller than remaining teeth, two paralateral teeth and 6-7 small lateral serrations; hypostoma with 1 + 1 line of five lateral setae and 1 + 1 short setae parallel to lateral margin and 1 + 1 or 2 + 2 short setae in posterior region of hypostoma near hypostomal groove. Subesophageal ganglion not pigmented (Fig. 37). Antenna as long as labral fan stalk; antennal articles pale brown except distal article, which is darker than proximal and medial articles (Fig. 39); segment proportions (proximal: medial: distal) 0.9-1.0: 1.0-1.1: 3.0-3.1 (n = 2). Mandible with three apical teeth, 17 internal teeth, one large mandibular serration, one small mandibular sensillum (sensu Craig & Craig 1986) and two lateral mandibular processes thick and longer than mandible margin (Fig. 42). Labral fan with 45-50 rays. Thorax gray dorsally and ventrally. Gill histoblast with six filaments. Abdomen usually completely yellowish gray dorsally, progressively paler ventrally, whitish posteroventrally; intersegmental bands faintly visible dorsally (Figs 40, 41). Ventral nerve cord dark gray. Ventral tubercles small, sub-triangular. Anterodorsal arms of anal sclerite shorter in length than posteroventral arms (Fig. 43). Posterior proleg bearing approximately 64 rows of up to 15-16 hooks (n = 2). Rectal papillae (Fig. 44) with three lobes, each with approximately 12-13 small finger-shaped lobules (n = 2). Etymology - The name of this species refers to the place where the holotype was collected - Pico dos Marins (Marins Peak) - a high mountain with granite outcrops in Brazil. Distribution - S. marins n. sp. is known from two localities - a small headwater stream on Pico dos Marins (Figs 1, 2), SP, and a small headwater stream in Serra dos Órgãos National Park, RJ, suggesting that this species is relatively widely distributed in high mountains. Bionomics - Larvae and pupae were collected in low numbers from a small (0.2 m wide) rocky-bottomed first-order stream. The stream on Marins Peak is situated in an open area exposed to sunlight at 2,220 m above sea level. The water temperature at the time of collection was 18ºC, the conductivity was below 10 μS/cm and the pH was 6.5. Larvae and pupae were collected from trailing vegetation and submerged deciduous leaves. We also collected two pupal exuviae from one small rocky-bottomed first-order stream located in Serra dos Órgãos National Park. This stream is situated in a forested area at 1,580 m above sea level with some incidence of sunlight. The water temperature at the time of collection was 19ºC, the conductivity was below 10 μS/cm and the pH was 5.9. Nothing is known about the biting habits of adult females. Type data - Holotype (MZUSP): Brazil, SP (near the border of MG), Piquete County. Pico dos Marins (Marins Peak), small stream, 22º29'48.6''S 45º07'22.2''W, 14.vii.2008 (M Pepinelli, T Siqueira) - pinned male (reared), with pupal exuviae (in glycerine vial). Paratypes: Brazil, SP (near the border with MG), Piquete County. Pico dos Marins (Marins Peak), small stream, 22º29'48.6''S 45º07'22.2''W, 14.vii.2008 (M Pepinelli, T Siqueira) - five larvae and one pupa in alcohol (MZUSP), one pinned female (reared) with pupal exuvia in glycerine (MZUSP), one pinned male (thorax) (reared) with head, genitalia and pupal exuvia mounted on slide (MZUSP), one pinned female (reared) with head, genitalia and pupal exuvia mounted on slide (MZUSP), five larvae in alcohol (INPA). Pico dos Marins (Marins Peak), small stream, 22º29'48.6''S 45º07'22.2''W, 25.viii.2005 (Roque FO) - two pupal exuviae mounted on slides (INPA). Brazil, RJ, Teresópolis County. Serra dos Órgãos National Park, small stream, 22º27'01.8''S 43º00'49.8''W, 18.vii.2008 (M Pepinelli, T Siqueira) - two pupal exuviae in alcohol (MZUSP). DISCUSSION S. marins n. sp. is assigned to the subgenus S. (Inaequalium) based on the diagnostic characters of Coscarón and Wygodzinsky (1984). The adults of most species of S. (Inaequalium) are morphologically similar, while the pupae are the best stage to distinguish the species, mainly the configuration of the pupal gill filaments (Coscarón 1987, Hernández et al. 2007). The female of S. marins n. sp. is similar to that of Simulium margaritatum Pepinelli, Hamada & Luz and Simulium petropoliense Coscarón by the shared absence of a pattern on the scutum (Fig. 4). However, the scutum of S. marins n. sp. is less densely haired than that of S. margaritatum and more densely haired than that of S. petropoliense (Pepinelli et al. 2006, Hernández et al. 2006, 2007). The anal lobe (= paraproct) of S. marins n. sp. female is sub-triangular with a small protuberance near the cercus (Fig. 15), similar to that of females of Simulium botulibranchium Lutz, S. petropoliense and Simulium souzalopesi Coscarón but, in the new species, the anal lobe is covered by microtrichia (Fig. 15). The male of S. marins n. sp. is similar to those of several other species of S. (Inaequalium) in the shape of the genitalia and by having the scutum covered in golden hairs without pattern. In the pupal stage, the new species is distinguished from all other nominal S. (Inaequalium) species by the combination of having six filiform gill filaments, a long anterodorsal projection on the cocoon and having the cephalic and thoracic plates covered with spines (Figs 27-34). The larva of S. marins n. sp. has a postgenal cleft in the shape of a narrow, semi-rounded, notch (Fig. 37), which distinguish it from the larvae of all other species of S. (Inaequalium). Two species groups (the inaequale-group and the botulibranchium-group) are currently recognized in S. (Inaequalium) (Coscarón 1987, 1991, Coscarón & Coscarón-Arias 2007, Hernández et al. 2007). However, the new species shares characters with both species groups. The sub-quadrangular gonocoxite and subcylindrical gonostylus of males, the subtriangular anal lobe of females and the small postgenal cleft of larvae are all characteristic of the botulibranchium-group. However, the slender (i.e., unswollen) and regularly configured gill filaments of pupae are characteristic of the inaequale-group. We suggest that features of the pupa should not be used to distinguish members of the two species groups because the gill filaments of S. (Inaequalium) species exhibit marked variation in shape and configuration, as illustrated by Hernández et al. (2007). Furthermore, Simulium rappae Py-Daniel & Coscarón and Simulium travassosi Lutz have swollen gill filaments but are currently assigned to the inaequale-group based on the morphology of male and female genitalia. In addition, males of S. marins n. sp. share with the other three species in the botulibranchium-group an elongated gonostylus, while species in the inaequale-group have a shorter and wider gonostylus (Hernández et al. 2007). Character states of the male and female strongly suggest that S. marins n. sp. should be assigned to the botulibranchium-group. S. marins n. sp. has the basal section of radius bare. Only two other species in the subgenus S. (Inaequalium)share this character: S. margaritatum and Simulium lundi (Py-Daniel & Barbosa). Although, Hernández et al. (2007) stated that females of S. margaritatum from state of Goiás (GO) have setae on the basal section of R, we do not agree with this assertion. S. margaritatum was described from populations collected in state of Bahia (BA) and all specimens examined lacked setae on the basal section of R (Pepinelli et al. 2006). We believe that there was a misidentification of the specimens from GO (Hernández et al. 2007) and that the specimens identified as S. margaritatum by those authors is a variant of S. rappae, a species that occurs in GO. We arrived at this conclusion by examining figures in Hernández et al. (2007) and by the fact that S. rappae has highly polymorphic gill filaments in terms of thickness and filament number. Illustrated specimens of S. margaritatum pupal gill filaments included in Hernández et al. (2007; Fig. 217) do not match those of S. margaritatum from the original description (Pepinelli et al. 2006; Figs 27-31); the gills in the original description are thinner and longer than those in populations from GO (Hernández et al. 2007). Another incongruence between S. margaritatum from BA (original description) and those from GO is the presence of a clear scutal pattern on females from the latter state (Hernández et al. 2007). Pepinelli et al. (2006) analyzed several females of S. margaritatum reared from pupae collected at different localities in BA and all of them had no patterns on the scutum. Another hypothesis is that the population from GO reported as S. margaritatum indeed represents a new species, with adults morphologically similar to those of S. rappae and pupae with four, thick, gill filaments. Additional studies including chromosomal analyses and molecular markers are needed to clarify the taxonomic status of GO population. ACKNOWLEDGMENTS To Dr. Fabio de Oliveira Roque, who collected this new species for the first time, and to Tadeu de Siqueira Barros, Reinaldo and people from Tribo Montanhismo (Orlando and Juliano, Itajubá, MG), who helped us during the field work. REFERENCES
Copyright 2009 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc09163f35.jpg] [oc09163f27-34.jpg] [oc09163f17-18.jpg] [oc09163f5-16.jpg] [oc09163f1-2.jpg] [oc09163f3-4.jpg] [oc09163f19-26.jpg] [oc09163f36-44.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}