 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 104, No. 5, August, 2009, pp. 790-796 SHORT COMMUNICATIONS Distribution of Lymnaeidae (Mollusca: Pulmonata), intermediate snail hosts of Fasciola hepatica in Venezuela JP PointierI; O NoyaII; B Alarcón de NoyaII; A ThéronI, + IUMR
5244 CNRS-EPHE-UPVD, Université de Perpignan, Via Domitia, 52 avenue
Paul Alduy, 66860 Perpignan Cedex, France Financial support: ECOS-Nord (V06A01), Instituto de Altos Estudios en Salud Pública Dr. Arnoldo Gabaldón Received 9 January
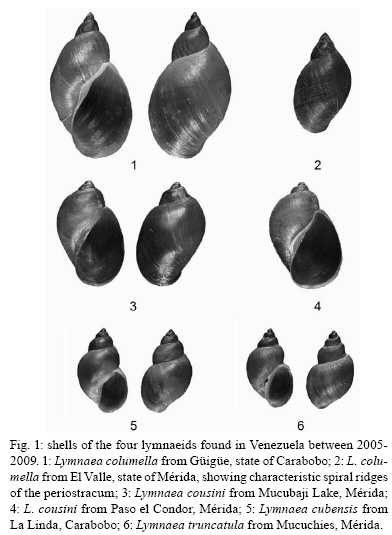
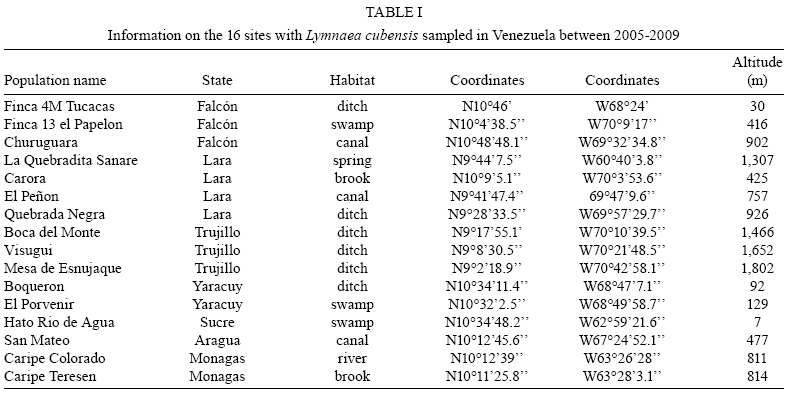
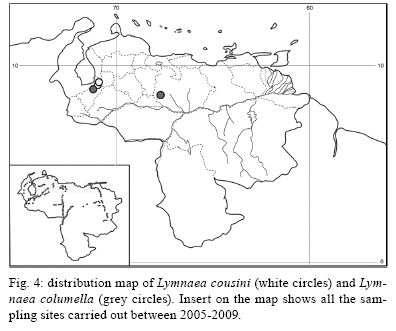
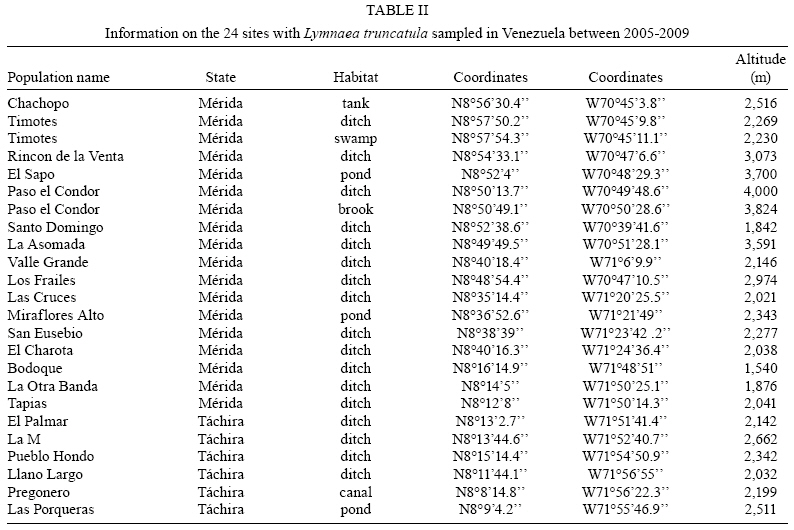
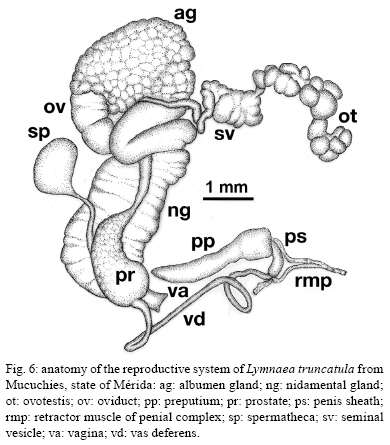
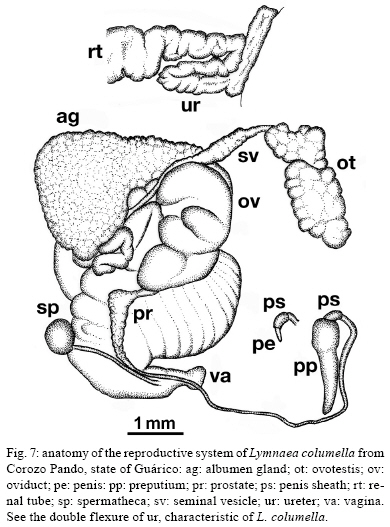
2009 Code Number: oc09173 ABSTRACT An extensive malacological survey was carried out between 2005-2009 in order to clarify the exact number of lymnaeid species which may be intermediate hosts of Fasciola hepatica in Venezuela. Four species were discovered during this survey, including two local species: Lymnaea cubensis and Lymnaea cousini and two exotic species: Lymnaea truncatula and Lymnaea columella. The most common local species was L. cubensis which was found at 16 out of the 298 sampling sites. This species has a large distribution area throughout the Northern part of Venezuela and was encountered from sea level to an altitude of 1,802 m in state of Trujillo. The second local species L. cousini was collected at only two sites of the Andean Region at altitudes of 3,550 m and 4,040 m, respectively. The European L. truncatula was found at 24 sites all located in the states of Mérida and Táchira at an altitude varying between 1,540-4,000 m. The respective distribution areas of L. cubensis and L. truncatula do not appear to overlap, but more detailed malacological surveys are needed. The fourth lymnaeid species, L. columella was collected in a canal from Mérida at an altitude of 1,929 m and in an irrigation canal from the state of Guárico, at an altitude of 63 m. The role of these four lymnaeid species in the transmission of fascioliasis in Venezuela is discussed. Key words: Lymnaeidae - Venezuela - distribution - fascioliasis Fascioliasis is a parasitosis mainly infecting cattle, but it is now considered to be an emergent disease in humans in many countries over the world (Mas-Coma 2005). In the New World, it is considered as a serious health problem in several Andean countries. Bolivia is even described as a hyperendemic area and one of its epidemiological characteristics is its very high altitude at more than 4,000 m (Esteban et al. 1999, Mas-Coma et al. 1999). The timing of the emergence of human foci in the Altiplano Region corresponds to the invasion by the European intermediate snail host Lymnaea truncatula (Jabbour-Zahab et al. 1997, Meunier et al. 2001). In Venezuela, human fascioliasis was first reported in 1910 and only eight sporadic cases were detected in the following decades (Risquez 1929 , Rodríguez & González 1975, Abdul-Hadi et al. 1996, Scorza et al. 1999, Incani et al. 2002, Alarcón de Noya et al. 2006). However, in 2005 five children belonging to the same family were detected with fascioliasis in Timotes, in the Venezuelan Andes, at an altitude of 2,230 m (Alarcón de Noya et al. 2007). The same year a malacological survey carried out in the Timotes area found only a single lymnaeid species, the European L. truncatula. In this paper we present the results of an extensive malacological survey which was carried out between 2005-2009 in order to clarify the exact number of lymnaeid species which may be intermediate hosts of Fasciola hepatica in Venezuela and to determine their current distribution throughout the country. A malacological survey was carried out between 2005-2009 across the whole country. A total of 298 sites were sampled (see the inserts in the maps showing all the sampling sites). Snails were sampled from different vegetation either by hand or sampled using a scoop mounted with a wooden handle, depending on the type of site. These sites include the following main habitat types: springs, ditches, brooks, canals, rivers, swamps, tanks, ponds and lakes. Following field collection lymnaeid snails were allowed to relax overnight using menthol. They were then immersed for 40 s in water at 70ºC, from which they were transferred to water at RT. The soft parts were drawn from the shell with small forceps and fixed in slightly modified Railliet-Henry fluid (distilled water 930 mL, sodium chloride 6 g, formalin 50 mL, glacial acetic acid 20 mL). Shells were measured to the nearest 1 mm using callipers. Snails preserved in Railliet Henry's fluid were dissected under a stereoscopic microscope and drawings of the reproductive system were made using a camera lucida attachment. Snails were identified according to conchological and anatomical characteristics (Paraense 1976, 1982, 1984, 1995, Brown 1994, Pointier et al. 2004, 2007). Four lymnaeid species were discovered during this survey in Venezuela, including two local species, Lymnaea cubensis L. Pfeiffer, 1839 and Lymnaea cousini Jousseaume, 1887 and two exotic species L. truncatula (Müller, 1774) and Lymnaea columella Say, 1817 (Fig. 1). The most common local species was L. cubensis which was found at 16 out of the 298 sampling sites. These 16 sites include six ditches, three canals, three swamps, two brooks, one river and one spring (Table I). L. cubensis has a large distribution area throughout the northern part of Venezuela from the oriental states of Sucre and Monagas to the occidental state of Trujillo (Fig. 2). This lymnaeid snail was encountered from sea level (Hato Rio de Agua, state of Sucre) to an altitude of 1,802 m in Trujillo (Mesa de Esnujaque) (Table I). Adult shells measured 6-10 mm and showed a large amount of variability. The anatomy of the reproductive system has two main reliable characters (Samadi et al. 2000): (i) small and ovoid-shaped prostate gland and (ii) a penis sheath about as long or smaller than the preputium (Fig. 3). A second local lymnaeid species L. cousini was reported for the first time in 2003 in Mucubaji Lake, state of Mérida, at an altitude of 3,550 m (N8º47'51.8'' W70º49'32.4'') (Pointier et al. 2004). Three years later in 2006 this species was discovered in a small ditch in the Paso el Condor area, Mérida, at an altitude of 4,040 m (N8º50'38.2'' W70º49'33.9'') (Fig. 4). Adult shells of L. cousini vary between 6.6-9.3 mm in heigth and have a large aperture and inflated body whorls separated by deep sutures (Fig. 1). These sizes are smaller than those from the original description of L. cousini by Jousseaume (1887) but the qualitative shell characters agree perfectly with those originally described. The anatomy of the reproductive system of the Venezuelan specimens is similar to that described by Paraense (1995) for topotypic specimens from Ecuador and has reliable characters: (i) a renal tube showing two distinct flexures of the ureter; (ii) a vagina with a bulbous appearance due to local thickening; (iii) a flat spermiduct with a granular surface; (iv) a prostate with the same granular appearance and a fissure formed by the folding of its left margin and (v) a penis sheath a little longer than the preputium (Fig. 5). A third lymnaeid species, the European L. truncatula found at 24 sites all located in Mérida and Táchira at an altitude varying between 1,540-4,000 m (Table II). This lymnaeid snail was found in 17 ditches, three ponds, one artificial tank, one brook, one swamp and one canal (Table II). Shells of L. truncatula are indistinguishable from those of L. cubensis (Samadi et al. 2000) (Fig. 1). The most relevant anatomical characters are the size and shape of the prostate gland and the relative lengths of the penis sheath and preputium: in L. truncatula, the prostate is much bigger and pear-shaped and the penis sheath is much shorter than the preputium (Fig. 6); in L. cubensis, the prostate is smaller and ovoid and the penis sheath a little shorter or about the same length as the preputium (Fig. 3). During this malacological survey, a fourth species, the exotic L. columella was also collected in a canal at El Valle, Mérida, at an altitude of 1,929 m (N8º38'40.3'' W71º7'28'') (Fig. 4) and in an irrigation canal at Corozo Pando, state of Guárico at an altitude of 63 m (N8º37'2.4'' W67º33'56.8'') (Fig. 4). Shells of the Vene-zuelan L. columella showed all the characteristics described by Paraense (1983) and Brown (1994): ovate shell with a short spire and pointed apex, a large last whorl about three times the length of the rest of the shell and the presence of characteristic minute spiral ridges of the periostracum (Fig. 1A, B). The most prominent anatomical features of L. columella are: (i) presence of a double flexure of the ureter; (ii) a thread-like to ribbon-like prostate; (iii) a preputium about 2-6 times as long as the penial sheath and (iv) a penial sheath apparently devoid of minute apical chambers (Fig. 7). Morphological identification of lymnaeids The members of the Lymnaeidae family exhibit a vast conchological diversity linked to substantial eco-phenotypic plasticity making a clear distinction between species difficult. This difficulty may explain why, until recently, only two species, L. cubensis and L. columella, have been reported in Venezuela (Lutz 1928, Briceño-Rossi 1950, Malek & Chrosciechowski 1964, Pino & Morales 1982, Morales & Pino 1992). However, the recent discovery of a third species, L. cousini at very high altitudes in the Venezuelan Andes, is perhaps linked to the scarcity of malacological data from this area rather than an identification problem, as this L. cousini has a shell quite distinct from the other species (Pointier et al. 2004). The use of anatomical characters of the reproductive system has proved very useful for the distinction between lymnaeid species in the Neotropical Area (Paraense 1976, 1982, 1984, 1995). These characters are particularly useful in separating the local L. cubensis from the European L. truncatula because conchological studies carried out on several populations of these two species, were unable to distinguish between them due to the large variability within and between populations (Samadi et al. 2000). In contrast, this study was able to clearly distinguish between the two taxa using characters of the male reproductive system. The validity of these anatomical markers has been confirmed by a genetic variability analysis based on 12 enzyme loci (Durand et al. 2002) and by the analysis of several DNA sequences such as the small subunit (18S) gene, internal transcribed spacers (1 and 2) and the cytochrome c oxidase subunit I (Bargues et al. 2007). Distribution of the lymnaeid snails in Venezuela The most prominent feature of our malacological survey is the discovery of 24 populations of the European L. truncatula in the Andinean Mérida and Táchira at an altitude varying between 1,540 m (Bodoque) and 4,000 m (Paso el Condor). These results confirm the invasion of L. truncatula into South America by which is now present in the Venezuelan Andes, the Altiplano Region of Bolivia and Peru (Mas-Coma 2005), the Argentinian Andes (Bargues et al. 2006) and the Valdivia area in Southern Chile (Yahia 1997). In Venezuela, the local species L. cubensis is sporadically distributed across the whole northern part of the country from the oriental state of Sucre to the western of Trujillo in the Andes where the species reaches an altitude of 1,802m at Mesa de Esnujaque. This last site is only about 10 km from Timotes, where L. truncatula was collected, but belongs to a different valley system. The respective distribution areas of L. cubensis and L. truncatula do not appear to overlap, but more detailed malacological surveys remain to be done in the Venezuelan Andes in order to see if a phenomenon of competitive exclusion occurs between the two species. In Venezuela, L. cousini was collected at only two sites at a very high altitude (3,550 m and 4,040 m, respectively). This discovery confirms that this species is restricted to high altitude habitats in the northern part of the Andean chain. Currently L. cousini has only been reported from ponds of the type locality in Ecuador at a height of 2,950 m (Jousseaume 1887) or from several sites in Ubaque and Bogotá, Colombia, at heights between 2,066-2,650 m (Piaget 1912, Pilbry 1935, Velásquez 2006). The two Venezuelan sites harboured monospecific populations of L. cousini. In this region, the rarity of L. cousini coupled with the occurrence of a number of similar sites colonized by L. truncatula, could be interpretated as the result of competitive displacement due to the invasion by L. truncatula, but this hypothesis remains to be investigated. The timing and origin of the introduction of L. truncatula in Venezuela are not known but molecular studies carried out on these lymnaeid populations may provide further information. Indeed, a study of the genetic diversity and population structure of L. truncatula samples collected from the Bolivian Altiplano has shown the presence of a single genotype. Several hypotheses have been discussed to explain this remarkable result and the most likely is that this species could have recently invaded the Bolivian Altiplano from Europe or from other areas of South America (Meunier et al. 2001). This hypothesis is supported by previous malacological surveys carried out in this area: the presence of lymnaeid snails on the Bolivian Altiplano was first reported by Ueno et al. (1975) and before that, lymnaeid snails were always absent from species lists made during surveys, the last being the Percy Sladen Trust Expedition of 1937 (Haas 1955). A second exotic lymnaeid species, L. columella, presently occurs in Venezuela. This species was first reported in a canal in the city of Maracay, state of Aragua (Malek & Chrosciechowski 1964) and its presence was confirmed at this site in 1996, as well in two other sites from the southern part of Valencia Lake, state of Carabobo (JP Pointer , O Noya, unpublished observations). However, recent surveys carried out in 2006 did not record L. columella from these sites. It seems that this species is now sporadically distributed in Venezuela and has been found from sea-level to high altitude, i.e., 1,929 m at El Valle, Mérida, in 2006. L. columella probably originates from North America, but now has a worldwide distribution. During the XIX century, its distribution in the United States extended over the eastern and midwestern states as well as from Canada to the southern states (Baker 1928). At the beginning of the XX century, it was introduced to California and Oregon (Vanatta 1915, Greg 1923). It was then reported from Cuba and Mexico (Aguayo 1938) and enlarged its distribution area to Puerto Rico (Harry & Hubendick 1964), Colombia (Malek & Cogswell 1980), Brazil (Paraense 1982) and Argentina (Scott 1953). This snail was also introduced to Australia (Boray et al. 1985) and to several Pacific islands, such as Tahiti, Rapa, Tubuai and Rurutu (Pointier & Marquet 1990, Gargominy & Fontaine, personal communication). In 1942 it was first reported from South Africa and then from several other African countries (Appleton 2003). The presence of this species in Europe had been restricted to botanical gardens (Brown 1994) but it was recently discovered in the wild in Southern France (Pointier et al. 2007). Role of lymnaeid snails in the transmission of fascioliasis in Venezuela The four lymnaeid species discovered at the present time in Venezuela are all susceptible to F. hepatica miracidial infection and their role as effective intermediate hosts has been demonstrated in several countries (Malek 1985). L. cousini has recently been found naturally parasitized by F. hepatica in Machachi, Ecuador (Villavicencio & Carvalho de Vasconcellos 2005), and in Paipa, Colombia (Velásquez 2006). However, as stated by Mas-Coma (2007), its role in the transmission of F. hepatica is well known in Colombia from long ago (Brumpt et al. 1939-1940). Unfortunately, the role of L. cousini as an intermediate host of fascioliasis in Venezuela is not known but its apparent rarity suggests it has a negligible epidemiological importance. The epidemiological situation seems similar to that of the introduced L. columella which is uncommon at the present time in Venezuela and which has never been reported naturally infected by F. hepatica in the field. However, all Venezuelan samples collected in this study are morphologically similar to susceptible phenotypes studied in Cuba, i.e., well rounded shell and characteristic mantle pigmentation pattern (Gutiérrez et al. 2003). Moreover, L. columella has been reported naturally infected by F. hepatica in several countries it has invaded, such as Australia (Boray et al. 1985), Brazil (Ueta 1980) and Argentina (Prepelitchi et al. 2003). In Venezuela, fascioliasis is a serious problem for cattle and has been reported from several states such as Mérida (Vivas 1976), Trujillo (Morales & Pino 1981), Lara, Yaracuy, Falcón, Portuguesa (Meléndez et al. 1983) and Zulia (Moreno & España 1982). In all cases, L. cubensis was reported as the intermediate snail host transmitting the parasite to cattle. This study on the distribution of L. cubensis in Venezuela agrees with all these observations (Fig. 2), except for the highest parts of Mérida, where the most common lymnaeid snail is L. truncatula and not L. cubensis. L. truncatula is the main snail host of human fascioliasis in the Old World and is responsible for the high prevalence and intensities which have been observed in humans in the Northern Bolivian Altiplano (Mas-Coma et al. 1999). In Venezuela, the occurrence of L. truncatula in the Timotes area where five positive children belonging to the same family were detected must be also emphasized (Alarcón de Noya et al. 2007). Consequently, the current presence of numerous L. truncatula populations in the Venezuelan Andes must be now considered as a new potential risk not only for cattle, but also for humans. Epidemiological implications of the presence of L. truncatula for cattle and humans living in this area must be investigated in the future. ACKNOWLEDGEMENTS To Dr. Suzanne Mills, for helpful revision of the manuscript. REFERENCES
Copyright 2009 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc09173t2.jpg] [oc09173t1.jpg] [oc09173f6.jpg] [oc09173f4.jpg] [oc09173f1.jpg] [oc09173f7.jpg] [oc09173f3.jpg] [oc09173f2.jpg] [oc09173f5.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}