 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 104, No. 6, September, 2009, pp. 862-864 ARTICLES Primary culture of intestinal epithelial cells as a potential model for Toxoplasma gondii enteric cycle studies Marcos de Assis MouraI; Maria Regina Reis AmendoeiraII; Helene Santos BarbosaI, + ILaboratório
de Biologia Estrutural Financial support: CNPq, CAPES, PAPES-Fiocruz, FAPERJ Received 23 June
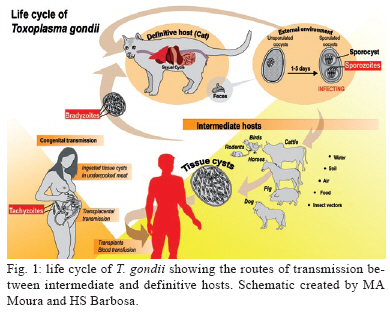
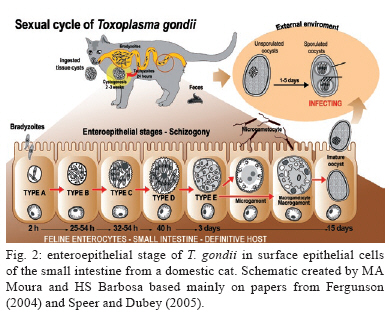
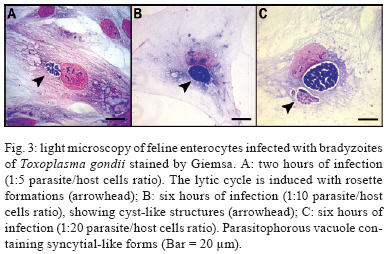
2009 Code Number: oc09182 ABSTRACT The primary culture of intestinal epithelial cells from domestic cats is an efficient cellular model to study the enteric cycle of Toxoplasma gondii in a definitive host. The parasite-host cell ratio can be pointed out as a decisive factor that determines the intracellular fate of bradyzoites forms. The development of the syncytial-like forms of T. gondii was observed using the 1:20 bradyzoite-host cell ratio, resulting in similar forms described in in vivo systems. This alternative study potentially opens up the field for investigation into the molecular aspects of this interaction. This can contribute to the development of new strategies for intervention of a main route by which toxoplasmosis spreads. Key words: Toxoplasma gondii - cat enteric cycle of T. gondii - intestinal epithelial cells - primary culture of enterocytes Toxoplasma gondii, the aetiological agent of toxoplasmosis, is an obligatory intracellular parasite, one of the most common zoonoses in the world. The wide distribution of the parasite is due to its multiple mechanisms of transmission: ingestion of cysts present in raw or poorly cooked meat; ingestion of oocysts in faeces of Felidae that contaminate food, water and soil (Dubey & Shen 1991, Uchoa et al. 1999, Dubey et al. 2004); by tachyzoite infection through vertical transmission during ingestion of milk in a breast-fed infant whose mother acquired a primary infection with T. gondii or horizontal transmission by ingestion of any type of milk which is a potential source of infection, in particular if consumed raw (Tenter 2009). The transmission cycle is demonstrated in Fig. 1. Approximately 25% of the human population is chronically infected (Black & Boothroyd 2000). The infection has clinical importance in two situations: (i) in congenital toxoplasmosis, where it can cause severe damage to the embryo in the form of mental retardation and blindness and (ii) as an opportunist infection in immunocompromised patients, inducing encephalic toxoplasmosis, which can be fatal (Dubey 2002, Hill & Dubey, 2002). On the 100th anniversary of the discovery of the parasite T. gondii, the literature presents a large number of scientific articles addressing such topics as toxoplasmosis (16,306 articles), T. gondii (9,659 articles) and infective stages: tachyzoite (1,106 articles), bradyzoite (262 articles) and oocysts (493 articles). Note, however, that the specific knowledge of the enteric cycle of T. gondii in Felidae is limited to 17 articles and is restricted to four articles in relation to the morphological characterisation in vivo (Frenkel et al. 1970, Dubey & Frenkel 1972, Fergunson 2004, Speer & Dubey 2005). The lack of cellular models that allow the reproduction of the enteric cycle of T. gondii exclusive of the definitive host has contributed to this picture. An alternative model to study the sexual cycle of T. gondii is the introduction of primary cultures of cat enterocytes. After ingestion of the tissue cysts by the definite host, the cyst wall is destroyed by proteolytic enzymes of the stomach and intestine, releasing the bradyzoites, which then penetrate the intestinal cells and begin the development of some generations of T. gondii (Dubey & Frenkel 1972). Five distinct morphological types, enteroepithelial stages or schizonts of T. gondii (types A, B, C, D and E) occur only in felines, the definite hosts, and involve the schizogony, gametogony and sporogony processes, resulting in the formation of immature oocysts (Dubey & Frenkel 1972, 1973, Dubey 1973, Dubey et al. 2004). The schizonts multiply exclusively in the superficial enterocytes of the small gut, giving origin to the merozoites, before the beginning of gametogenesis (Dubey & Frenkel 1972, Frenkel & Dubey 1972). The microgametes are then liberated in the small gut and are attracted to the macrogamonts. The immature oocyst is formed in the intestinal epithelium of felines and eliminated, still immature, together with faeces into the environment. The mature oocysts are formed by sporogony, a process that occurs outside the host and leads to the development of infectious oocysts, which contain two sporocysts, each containing four sporozoites (Dubey & Frenkel 1973) (Fig. 2). The morphologic and molecular characterisation of the enteroepithelial stages of T. gondii, including the sexual cycle, described thus far was realised in in vivo studies, using domestic feline neonates (Dubey & Frenkel 1972, Ferguson et al. 1974, 1975, Koyama et al. 2000, Ferguson 2004, Speer & Dubey 2005). The present study is a pioneering work showing the development of T. gondii schizonts in vitro similar to what has been previously described in vivo. Primary cultures of feline intestinal epithelial cells were obtained from the foetus of a clinically healthy pregnant domestic cat (no gastrointestinal disease and serologically negative for T. gondii, feline immunodeficiency virus and feline leukaemia virus). The samples of small intestine were collected aseptically corresponding to the jejunum-ileum region. The fragments were placed in nonenzymatic dissociation buffer (pH 7.2) containing 1mM EDTA, 1mM EGTA, 0.5 mM dithiothreitol and 10% antibiotic solution for 20 min under agitation at RT (Perreault & Jean-Francois 1996, Macartney et al. 2000, Aldhous et al. 2001, Rusu et al. 2005). The cell aggregates were plated in DMEM/Hams medium [Dulbecco's Modified Eagle's Medium/Hams Nutrient F12 (1:1)] containing 10% antibiotic solution, 1 mM glutamine, 10% bovine foetal serum and 20 ng/mL epidermal growth factor (Sanderson et al. 1996, Aldhous et al. 2001). The cultures were incubated at 37°C in a 5% CO2 atmosphere and the medium was renewed every two days. The use of bradyzoites of T. gondii (Guimarães et al. 2008) to infect feline intestinal epithelial cells is justified because it represents the major route of parasite transmission, through the consumption of raw meat by animals or humans. Another prominent point of this paper is the introduction of feline cells for the first time as a cell model to study the T. gondii-host cell interaction. Our results point out that the intracellular fate of the parasite is dependent on the parasite-host cell ratio employing ME49 strain bradyzoites: (i) the development of the lytic cycle of the parasite, using the 1:5 parasite-host cell ratio, with conversion of bradyzoites in tachyzoites, culminating with their intense proliferation and rosette formations (Fig. 3A); (ii) maintenance and establishment of bradyzoites with the formation of cysts, using a 1:10 bradyzoite-host cell ratio (Fig. 3B) and (iii) the development of the syncytial-like forms of T. gondii (Fig. 3C), using using the ratio of 1:20 (bradyzoite per host cell), similar to those forms described in vivo. Our results also showed that the reduction of the 1:10 bradyzoite-host cell ratio leads to the spontaneous formation of intracellular cysts, without any physical, chemical or immunological modulation. These data introduce new biological aspects of T. gondii host cell interaction. The experimental strategies implemented in the present work reproduced in vitro the natural microenvironment established during enteric development of T. gondii in the definitive host, the domestic cat. The introduction of the feline epithelial intestinal cell (FEIC) as a cellular model shows that it can potentially contribute to new approaches to investigating the cell biology of the parasite. Also, FEIC is an alternative methodology to better understanding the enteric cycle of T. gondii under controlled conditions, opening up the field for investigation of the molecular aspects of this interaction and contributing, for example, to the development of new strategies aimed at intervention in one of the main routes by which toxoplasmosis spreads. ACKNOWLEDGMENTS To Marielle Delfin Pereira and Sandra Maria de Oliveira Souza, for their technical assistance, and to David Straker, for the English revision. REFERENCES
Copyright 2009 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc09182f3.jpg] [oc09182f2.jpg] [oc09182f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}