 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 104, No. 6, September, 2009, pp. 933-934 SHORT COMMUNICATIONS Larval recovery of Toxocara cati in experimentally infected Rattus norvegicus and analysis of the rat as potential reservoir for this ascarid Sérgio V SantosI, +; Susana Z LescanoI; João M CastroII; Pedro P ChieffiI, III IInstituto
de Medicina Tropical de São Paulo (LIM 06), Av. Dr. Enéas de Carvalho
Aguiar 500 2º andar, 05403-000 São Paulo, SP, Brasil Received 6 March
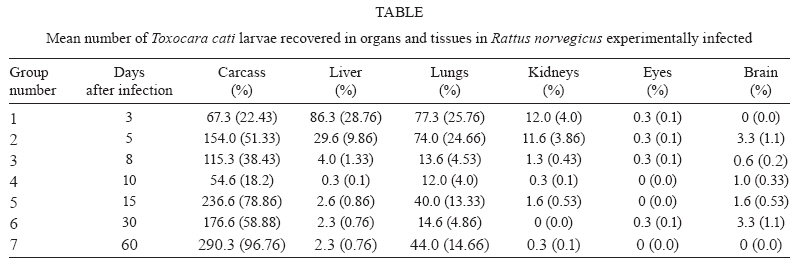
2009 Code Number: oc09195 ABSTRACT Toxocara cati is a common feline parasite transmitted by the ingestion of embryonated eggs, by the transmammary route or by predation of paratenic hosts harbouring third-stage larvae in their bodies. In the present study, the larval distribution of T. cati in tissues and organs of Rattus norvegicus experimentally infected with 300 embryonated eggs was analysed. Third-stage larvae were recovered from livers, lungs, kidneys, eyes, brains and carcasses of infected rats, following tissue digestion with HCl 0.5% for 24 h at 37°C. Some differences from the known larval distribution of Toxocara canisin the same rodent species were found. Key words: Toxocara cati - larval distribution - Rattus norvegicus - paratenic hosts Visceral larva migrans (VLM) is the result of prolonged tissue migration of nematode larvae in unusual hosts (Beaver et al. 1952). In human beings, the main etiological agents for this syndrome are either Toxocara canis or Toxocara cati larvae (Schantz 1989). T. cati is a common helminth parasite of cats and is closely related to T. canis, a roundworm that frequently infects dogs. When compared to the frequency of known human cases of infection by T. canis larvae, few records implicating T. cati as causative agents of VLM have been published (Fisher 2003). As a consequence, while T. canis is recognised as a dangerous zoonosis agent, T. cati has had its importance neglected by sanitary authorities and even sometimes by health professionals (Fisher 2003). However, there are some records of human intestinal infection by adult T. cati (Sprent 1956, Dubey 1968, Fisher 2003) and human ocular involvement by T. cati larvae causing ocular larva migrans (OLM) (Sakai et al. 1998), as well as some cases of VLM (Fisher 2003). Sprent (1956) pointed out that the larvae of T. cati can infect and migrate into tissues of some types of invertebrates (earthworms and cockroaches) as well as in birds, mice, lambs, dogs and probably human beings; the adult stage is only found in cats and, in rare cases, humans. Cats can become infected with T. cati by ingesting embryonated eggs found in contaminated soil or by predation of paratenic hosts - usually small rodents or birds containing live third-stage larvae in their tissues (Sprent 1956). Another means of transmission in cats is larval transmammary migration (Sprent 1956, Coat et al. 2004). Some behavioural changes were found in paratenic hosts harbouring third-stage T. canis larvae, probably favouring predation by dogs (Cox & Holland 2001), but there are no records of the occurrence of similar behavioural changes in rodents infected by T. cati. Rattus norvegicus has been recognised as a usual paratenic host for T. canis (Chieffi et al. 1981) and probably also for third-stage T. cati larvae. Although the larval distribution of T. canis in R. norvegicus has already been studied (Lescano et al. 2004), the larval distribution for T. cati has only been analysed in mice (Dubey 1968) and has shown some significant differences when compared to that found for T. canis in the same rodent (Bardón et al. 1994). The aim of the present study was to recover larvae from tissues and organs of experimentally infected R. norvegicus, in order to characterise the migration pattern of T. cati larvae in this rodent species. Twenty-one male R. norvegicus (Wistar strain) rats, aged eight weeks, were orally infected with 300 embryonated T. cati eggs containing third-stage larvae (Araújo 1972). On days 3, 5, 8, 10, 15, 30 and 60 after inoculation, three rats were killed and larvae were recovered from the liver, lungs, kidneys, eyes, brain and carcass of each rat after digestion with HCl 0.5% for 24 h at 37°C. Following sedimentation, supernatant was poured into a tube and centrifuged for 2 min at 1,500 rpm; then, 2 mL of the pelleted sediment was collected and thoroughly mixed and 0.1 cm3 samples were completely analysed under a light microscope for larval counts (Wang & Luo 1998). The final result, shown in Table, is an average of the larval numbers of all animals within each group. The experimental protocol was approved by the Research Ethical Committee of the Instituto de Medicina Tropical, São Paulo. Analysing T. canis larval migration, Lescano et al. (2004) observed a higher concentration of larvae in the liver, lungs and kidneys within the first five days after inoculation. After the 15th day of infection, an increase in larval recovery from the brain was noted, lasting up to the 30th day of observation. Larvae were found in the muscles from the fifty day of infection up to the end of the experiment. Some significant differences were observed in our study of R. norvegicus infected with T. cati larvae and the study of Lescano et al. (2004). In our study, for example, larval migration to the brain was less expressive - similar to that previously observed in mice (Bardón et al. 1994) - and occurred mainly on the 15th and the 30th days after inoculation. On the other hand, our report of the ocular migration of T. cati larvae occurring principally at the beginning of the this experiment confirms the findings of Akao et al. (2000) in experimentally infected gerbils and the hypothesis regarding the importance of T. cati larvae in the etiology of OLM. Under the conditions of our present experiment (a single inoculation of T. cati eggs and only 60 days of observation period), R. norvegicus does not seem to be the best model for OLM when compared to other rodent species (Havasiová-Rei-terová et al. 1995, Alba-Hurtado et al. 2009). However, it is important to highlight the raised level of R. norvegicus infection by Toxocara spp. in natural conditions (Chieffi et al. 1981), suggesting a role for this rat as a potential reservoir for Toxocara spp. in the environment. REFERENCES
Copyright 2009 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc09195t1.jpg] |
| |||||||||

{kind=link}