 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
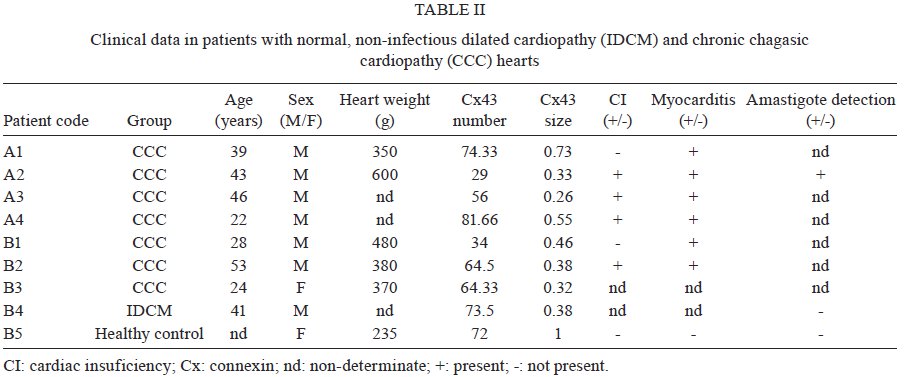
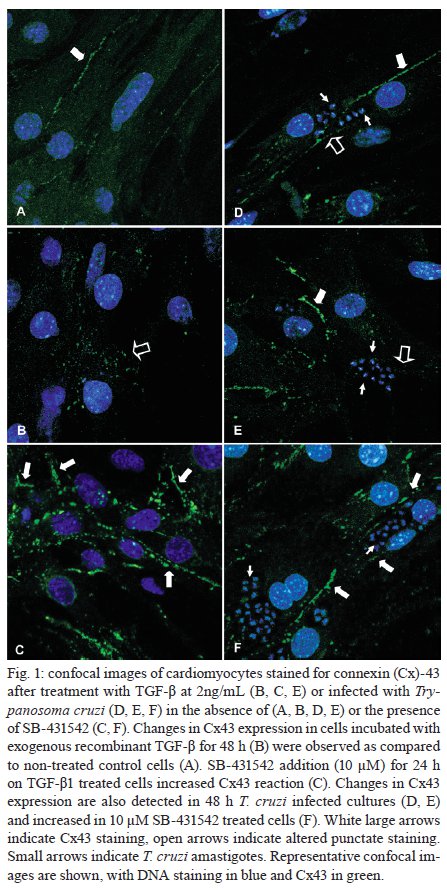
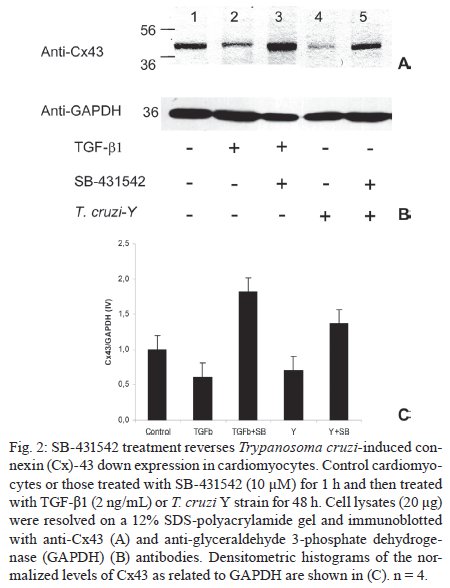
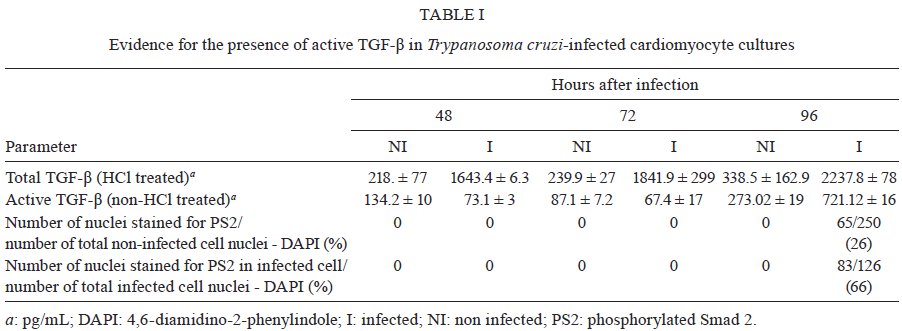
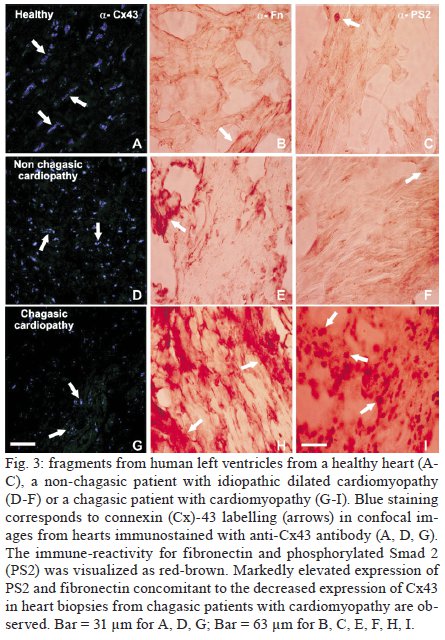
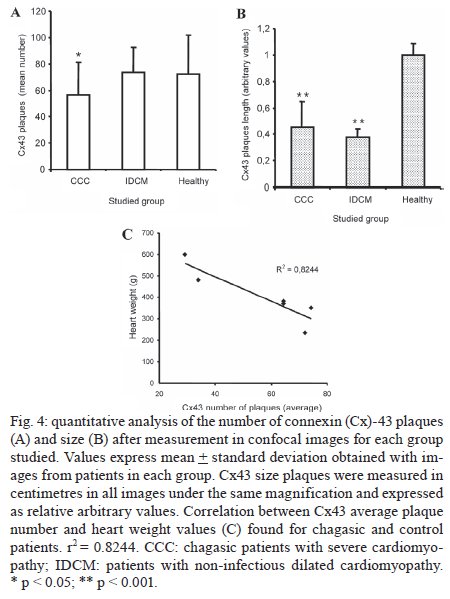
Memórias do Instituto Oswaldo Cruz, Vol. 104, No. 8, 2009, pp. 1083-1090 ARTICLES Gap junction reduction in cardiomyocytes following transforming growth factor-ß treatment and Trypanosoma cruzi infection Mariana C WaghabiI, II; Robson Coutinho-SilvaII, III; Jean-Jacques FeigeIV; Maria de Lourdes HiguchiV; David BeckerVI; Geoffrey BurnstockII; Tânia C de Araújo-JorgeI, + ILaboratório
de Inovações Terapêuticas, Instituto Oswaldo Cruz-Fiocruz,
Rio de Janeiro, RJ, Brasil Financial support: CNPq, FAPERJ, PRONEX, Wellcome Trust (062754/Z00Z to RCS) None of the authors have commercial or other associations that might pose a conflict of interest. Received 30 April 2009 Code Number: oc09222 ABSTRACT Gap junction connexin-43 (Cx43) molecules are responsible for electrical impulse conduction in the heart and are affected by transforming growth factor-β (TGF-β). This cytokine increases during Trypanosoma cruzi infection, modulating fibrosis and the parasite cell cycle. We studied Cx43 expression in cardiomyocytes exposed or not to TGF-β T. cruzi, or SB-431542, an inhibitor of TGF-β receptor type I (ALK-5). Cx43 expression was also examined in hearts with dilated cardiopathy from chronic Chagas disease patients, in which TGF-β signalling had been shown previously to be highly activated. We demonstrated that TGF-β treatment induced disorganised gap junctions in non-infected cardiomyocytes, leading to a punctate, diffuse and non-uniform Cx43 staining. A similar pattern was detected in T. cruzi-infected cardiomyocytes concomitant with high TGF-β secretion. Both results were reversed if the cells were incubated with SB-431542. Similar tests were performed using human chronic chagasic patients and we confirmed a down-regulation of Cx43 expression, an altered distribution of plaques in the heart and a significant reduction in the number and length of Cx43 plaques, which correlated negatively with cardiomegaly. We conclude that elevated TGF-β levels during T. cruzi infection promote heart fibrosis and disorganise gap junctions, possibly contributing to abnormal impulse conduction and arrhythmia that characterise severe cardiopathy in Chagas disease. Key words: gap junction - heart - Chagas disease - transforming growth factor-β - connexin 43 Gap junctions are aggregates (plaques) of intercellular channels that connect the cytoplasm of adjacent cells. They mediate both electrical and metabolic coupling. The gap channels are composed of 12 connexin (Cx) molecules assembled from the hexameric hemichannels (connexons) in humans (Sohl & Willecke 2004). In the heart, intercellular gap junction channels constructed from Cx molecules are crucial for propagation of electrical impulses in excitable tissues, most notably including myocardium, where Cx43 is the most important isoform and has been studied extensively (Severs 2001, Rodriguez-Sinovas et al. 2007). Some myocardial diseases show alterations in expression levels of Cxs and/or the anisotropic distribution of gap junction plaques (Severs et al. 2004, Chen & Zhang 2006). Decrease in expression levels of ventricular Cx43 has been described in the hypertrophic heart of hypertensive rats (Bastide et al. 1993) and in ischaemic human and rabbit hearts (Peters et al. 1993, Tansey et al. 2006). A redistribution of Cx43 gap junctions has been reported in the borderzone of infarcts (Severs et al. 2004, Chen & Zhang 2006), in hypertrophied hearts (Uzzaman et al. 2000), ischemic hearts (Peters et al. 1993, Beardslee et al. 2000) and hypertrophic cardiomyopathy (Sepp et al. 1996). The redistribution is frequently accompanied by a decrease in the size of gap junction plaques (Kaprielian et al. 1998) or in the total amount of plaques (Uzzaman et al. 2000). Some dysfunction in gap junction intercellular communication is regulated by growth factors [e.g., epithelial growth factor, transforming growth factor-β (TGF-β) and bovine pituitary extract], which may act to isolate affected cells from one another by down-regulating the transfer of ions and small molecules through gap junctions (Chanson et al. 2005). TGF-β is a cytokine that contributes to fibrosis by increasing the synthesis of collagen and other extracellular matrix proteins and decreasing their degradation. Physiological levels of TGF-β maintain tissue homeostasis and aberrant over-expression of TGF-β leads to tissue fibrosis (Tabibzadeh 2002). Through regulation of Cx43 expression, TGF-β affects gap junction-mediated intercellular communication (van Zoelen & Tertoolen 1991, Hurst et al. 1999, Rudkin et al. 1996) and the outcome varies between positive and inverse relationships, depending on the cell type, the type of membrane receptors employed and the initial activation/phosphorylation state of the cells (Chanson et al. 2005). Acute down-regulation of Cx43 at wound sites leads to a reduced inflammatory response, enhanced keratinocyte proliferation and wound fibroblast migration (Mori et al. 2006). Cardiomyopathy occurs in chronic Chagasic disease, which is an infection caused by Trypanosoma cruzi that is characterised by focal myocarditis following parasite invasion and inflammatory cell infiltration of heart myofibers (Palomino et al. 2000) that is likely to be triggered by parasite-host interaction during the initial acute phase. T. cruzi infection affects over 17 million people in Latin America, leading to 45,000 deaths/year (Schmunis 2007). However, a clear understanding of its pathophysiology remains a challenge (Marin-Neto et al. 2007). Most of the infected people remain asymptomatic in the so-called "indeterminate form"; however, about 25% of the patients develop the cardiac clinical form, wherein the heart increases due to exacerbated fibrosis, leading to dysfunctions such as arrhythmias, conduction disturbances and congestive heart failure (Higuchi et al. 1999). T. cruzi infection of cultured rat and mouse cardiac myocytes results in a decrease in electrical coupling between the infected cells, decreased passage of dyes into or away from infected cells and a decrease in the expression of organised plates of gap junction channels at appositional membrane areas (Campos de Carvalho et al. 1992, Adesse et al. 2008). We have demonstrated previously TGF-β involvement in heart pathology in mice following acute infection and in human chronic heart fibrosis occurring in Chagas disease (Araújo-Jorge et al. 2008). Herein we studied in vitro cultures and heart samples from chagasic patients, in which TGF-β signalling has previously been shown to be highly activated, to test the hypothesis that the expression of Cx43 would decrease under high levels of TGF-β (Araújo-Jorge et al. 2002). PATIENTS AND METHODS In vitro T. cruzi heart cell infection - Cardiomyocytes from mouse embryos were obtained as described previously (Meirelles et al. 1986). Cells were seeded in 24-well plates and maintained in Eagle's medium containing 0.1% fetal calf serum, garamicine, 1 mM glutamine and 2.5 mM CaCl2. To analyse the localisation of Cx43, some subconfluent monolayers were incubated with T. cruzi trypomastigotes from the Y strain, employing a parasite/host cell ratio of 10:1, along with TGF-β 1 (2 ng/mL) for 24 h at 37ºC. After washing, some monolayers were treated with SB-431542 (10 μM) for longer than 24 h, washed twice in phosphate buffered saline (PBS), fixed in 4% paraformaldehyde for 10 min at 4ºC and processed for immunocytochemistry analysis. SB-431542 is an activin receptor-like kinase (ALK)-5 inhibitor that blocks the TGF-β signalling pathway (Inman et al. 2002). All procedures were carried out in accordance with the guidelines established by the Fiocruz Committee of Ethical for the Use of Animals, resolution 242/99. Immunofluorescence and confocal microscopy of immunocytochemical staining - Cells were incubated once with 0.1% Triton X-100 in PBS for 10 min, followed by three 10 min washes in PBS/2% bovine serum albumin and then reacted with rabbit anti-Cx43 (Zymed, San Francisco, CA, USA) diluted 1:50 in PBS overnight at 4ºC. The monolayers were incubated for 1 h at RT with goat-anti-rabbit IgG-fluorescein isothiocyanate diluted 1:200 (Jackson ImmunoResearch Laboratories, West Grove, PA, USA). The cultures were then incubated with 4,6-diamidino-2-phenylindole (DAPI) (1:5000) to stain DNA, washed three times in PBS and mounted with CitiFluor AF1 (Agar Scientific Ltd, Stansted, Essex, UK). The sections were observed using a microscope Olympus Fluoview 3.3 (Olympus Corporation, Japan) coupled with an upright BX51 microscope image acquisition system. TGF-β measurement - Measurement was performed using a TGF-β 1-specific commercial enzyme linked immunosorbent assay (ELISA) kit (TGF-β 1 Emax; Promega Corporation, Madison, Wisconsin, USA) according to the manufacturer's instructions (the minimum detectable dose of TGF-β 1 in this assay is less than 32 pg mL-1). Supernatants of control cardiomyocytes or those infected with T. cruzi at 48, 72 and 96 h post-infection were assayed after acidic pH activation of latent TGF-β 1 (total TGF-β) or without activation (active TGF-β) to calculate the ratio between active and latent TGF-β 1. Immunoblot analysis - Cells were treated or not with SB-431542 (10 μM) for 1 h and then TGF-β 1 (2 ng/mL) or T. cruzi Y strain were added for 48 h. Cardiomyocytes were then washed twice with PBS and lysed in radioimmunoprecipitation assay buffer [50 mM Tris-HCl, NaCl 150 mM, 1% Triton X-100, 1 mM ethylene glycol-bis-(b-amino-ethyl ether) N,N,N´,N´-tetra-acetic acid, 100 μg/mL phenylmethylsulfonyl fluoride, 1 μg/mL pepstatin, 10 μg/mL leupeptin and 1 μg aprotinin, pH 8.0)]. Proteins in the lysates (20 μg/lane) were separated by sodium dodecyl sulfate polyacrylamide gel electrophoresis (12%) and analysed by immunoblotting with polyclonal rabbit anti-Cx43 (Sigma Aldrich, St. Louis, IL, USA) diluted 1:8000. The same membrane was stripped and re-probed with a monoclonal antibody against glyceraldehyde 3-phosphate dehydrogenase (GAPDH) to confirm equal protein loading in each lane. Four independent experiments were performed and the densitometric histograms of the normalised levels of Cx43 as related to GAPDH are shown. Heart cryosections - Left ventricle fragments from the heart of patients chronic chagasic cardiopathy (CCC) (n = 7) as well as from patients with non-infectious dilated cardiopathy (IDCM) (n = 1) and a reference control from a non-infected person (Healthy control) (n = 1) were obtained as described elsewhere (Araújo-Jorge et al. 2002). Hearts were taken at autopsy from four patients (Table II, A1-4) who had died from cardiac failure, including two men aged 39 and 43 years (A1, 4) who died from ventricular fibrillation leading to severe arrhythmia, a man aged 46 years (A2) who died from cardiogenic shock by ventricular failure, a young man aged 22 years (A3) and biopsies from four patients (Table II, B1-4) that underwent heart transplant (heart explanted from the recipients). Three hearts were obtained from a chagasic group (B1-3, 2 men aged 28 and 53 years, or class IV and a woman aged 24 years, NYHA class III) and one from a patient from the IDCM group (B4) with IDCM (NYHA class III, a man aged 41 years, presenting idiopathic dilated cardiopathy). The biopsy of one young woman who presented normal heart function, but was submitted to valve prosthesis, was used as a non-infected control (B5). Immunohistochemical staining - Processing for immunofluorescence was performed as described previously (Coutinho-Silva et al. 2001). Frozen sections (10 μm) were fixed for 10 min in 4% paraformaldehyde (VWR International, Poole, UK) in PBS followed by blocking with 10% normal horse serum (Harlan Sera-Lab, Loughborough, UK) in PBS containing 0.05% thimerosal (Merthiolate; Sigma Chemical Co, Poole, UK) for 20 min. The slides were then incubated overnight with mouse monoclonal antibodies against Cx43 (1:200) (Becker & Davies 1995). All incubations were held at RT and separated by three 5 min washes in PBS. For confocal microscopy, the secondary antibody was anti-mouse Cy5 diluted 1:200 in PBS for 1 h at 37°C, washed and mounted with glycerol CitiFluor AF1 (Agar Scientific). The sections were observed using a Zeiss Axioplan microscope (Zeiss, Oberkochen, Germany) coupled with a Leica DC 200 image acquisition system (Leica, Heerbrugg, Switzerland). Cx43 plaque quantification - Leica TCS NT confocal images were digitally processed using Photoshop 5.0 software (Adobe Systems, San Jose, CA, USA). Each test area was photographed at a final print magnification of 400X. At this magnification, a typical test area included approximately 34.600 μm2 of tissue area. The images were projected on a square screen and adjusted for 1 cm (arbitrary value) of length of Cx43 from healthy heart. The minimum gap junction length measured was 0.1 cm that corresponded to five contiguous pixels. The mean number of Cx43 plaques as well as their sizes was evaluated in each case studied by quantification of three confocal field images/patient. The means obtained in measurements from the IDCM and healthy patients were compared to the mean calculated using data collected from the seven chagasic cases. Analyses and tests were carried out using GraphPad Prism version 3.0 for Windows (GraphPad Software, San Diego, CA, USA). Statistical differences between the three groups were determined by analysis of variance followed by Dunnett's post-test. Statistical significance was defined as p < 0.05 and p < 0.001. Fibronectin and phosphorylated Smad 2 detection in heart cryosections - Cryostat slides were obtained as described above. Histology and immune labelling of T. cruzi antigens and of inflammatory infiltrates were already studied and described (Higuchi et al. 1997, Reis et al. 1997). Sections were fixed in paraformaldehyde, incubated with 1% H2O2, washed and saturated with 5% normal goat serum before an overnight incubation with rabbit IgG anti-fibronectin (Sigma) or anti-phosphosmad2 (PS2). The anti-PS2 antibody was raised in rabbits against the peptide KKKSSpMSp (where Sp stands for phosphorylated serine residues) and was given to us by Dr P ten Dijke (The Netherlands Cancer Institute, Amsterdam, The Netherlands). Its specificity was assessed previously by western blotting and immunochemical methods (Rosendahl et al. 2001). A biotin-avidin-peroxidase system (Dako, Glostrup, Denmark) was used for immune detection. The patients gave signed consent for their participation according to the rules of the official Fundação Oswaldo Cruz Ethical Committee (Brazilian Ministry of Health). RESULTS Expression of Cx43 in cardiomyocytes in vitro is reduced and disorganised after TGF-β treatment and after T. cruzi infection - To test for a biological effect of TGF-β on Cx43 expression, we treated in vitro cardiomyocytes with 2 ng/mL TGF-β for 48 h. In non-treated cultures, typical gap junction immunoreactivity was seen at virtually every interface between the cultured cardiomyocytes (Fig. 1A, arrow). A different, punctate and disorganised pattern in the Cx43 staining was observed following the addition of 2 ng/mL TGF-β (Fig. 1B, open arrow). This pattern indicated a clear effect of TGF-β on Cx43 expression and/or distribution. Western blot detection of Cx43 expression further confirmed the effect of TGF-β (Fig. 2A, C). Infection of cardiomyocytes with T. cruzi led to a decrease in Cx43 expression (Campos de Carvalho et al. 1992, Adesse et al. 2008) as detected by confocal microscopy and typical T. cruzi DNA staining by DAPI indicated the infected cells (Fig. 1D-F, small arrows). We confirmed the down-regulation of Cx43 staining in T. cruzi-infected cells (Fig. 1D, E, green labelling, open arrows) compared to non-infected cells (Fig. 1A, arrow). The cells containing parasites were negative for Cx43 staining (Fig. 1D, E, open arrow); however, non-infected cells from the same cultures sometimes retained the normal pattern of Cx43 expression (Fig. 1D, large arrow). Western blot detection of Cx43 confirmed its reduction after T. cruzi infection (Fig. 2A, Lane 4). SB-431542 treatment inhibits the effect of TGF-β on Cx43 expression in normal and T. cruzi-infected cardiomyocytes - Treatment of cardiomyocytes with SB-431542 strongly increased the Cx43 immunostaining (Fig. 1C), thus confirming that the TGF-β signalling effect did indeed specifically disorganise Cx43 expression in cardiomyocytes (Fig. 1B). Western blot analysis also showed that the addition of SB-431542 (10 μM) to non-infected TGF-β-treated cardiomyocytes up-regulated the expression of Cx43 (Fig. 2A, Lane 3). We recently demonstrated that SB-431542 inhibited T. cruzi infection (Waghabi et al. 2007), but in the present work we aimed to verify if the effect of SB-431542 was due to the blocked expression of TGF-β-responsive genes rather than a decreased infection rate. We then extended this approach by adding SB-431542 after 24 h of infection. We verified that treatment with SB-431542 of the 24 h-infected cultures (Fig. 1F) reversed the typical pattern of Cx43 reduction (Fig. 1D, E). Importantly, even infected cells expressed Cx43 (Fig. 1F, arrows). Western blot analysis confirmed that cardiomyocytes treated with SB-431542 (10 μM) prior to T. cruzi infection and/or TGF-β addition overexpressed Cx43 compared to the same procedure performed without SB-431542-mediated inhibition (Fig. 2A, Lanes 3, 5). TGF-β secretion by T. cruzi-infected cardiomyocytes - Previous reports have shown that cardiomyocytes secrete TGF-β in vitro (Flanders et al. 1995) and that Chagas disease is accompanied by increased TGF-β levels (Silva et al. 1991, Araújo-Jorge et al. 2002, Waghabi et al. 2002). We compared TGF-β levels measured in the supernatants of control cardiomyocytes and those infected by T. cruzi after 48, 72 and 96 h post-infection (Table I). Non-infected cells secreted regular levels of TGF-β over the period studied (70-250 pg/mL). Infected cardiomyocytes secreted higher TGF-β levels, starting from 1,600 and reaching 2,200 pg/mL, which is seven times higher than total TGF-β secretion from non-infected cells. Detection of active TGF-β in culture supernatants was similar in infected and non-infected cells after 48 and 72 h, but was 2.6-fold higher in the infected cells 96 h after infection. To confirm these observations, we performed immunostaining of cardiomyocytes infected by T. cruzi using an anti-TGF-β antibody. Amastigote-positive cells stained strongly with anti-TGF-β antibody (Waghabi et al. 2005). Non-infected cardiomyocytes surrounding infected ones were strongly immunoreactive for TGF-β by 24 h post-infection (Supplementary data A, arrow) as well as at 48 h (data not shown), 72 h (Supplementary data B) and 96 h (data not shown). These data confirmed TGF-β secretion by non-infected cardiomyocytes in infected cultures, which contributed to the available total and active amounts of this cytokine as measured by ELISA (Table I). We showed, by RT-PCR assays, that the addition of TGF-β to cardiomyocyte cultures increased by 3.6-fold the transcription of the TGF-β gene, thus demonstrating an autocrine/paracrine self-regulation (Supplementary data C). In addition, western blot analysis showed that Smad-2 phosphorylation following TGF-β addition to cardiomyocytes was decreased when incubated with SB-431542 in a concentration-dependent manner (Supplementary data D). Cx43, fibronectin and PS2 detection on human left ventricle of chagasic patients - Circulating levels of TGF-β and tissue expression of PS2 were associated with the progression of heart damage in Chagas disease (Araújo-Jorge et al. 2002). To compare and confirm the relevance of the in vitro data with human pathology, the expression and localisation of Cx43 was studied in the human left ventricle from healthy or cardiopathic patients. Confocal images showed staining for Cx43 (dark blue) in the different groups studied (Fig. 3A, D, G). In the healthy control sample, Cx43 labelling was localised at intercalated discs as a dense structure of plaques (Fig. 3A, arrows) as described previously (4). A change in Cx43 expression occurred in chagasic patients and we detected the disassembly of the gap junctions and a marked decreased in total staining (Fig. 3G). The mean number of Cx43 plaques was 20% lower in chagasic patients compared to healthy or IDCM groups (Table II, Fig. 4A). Cx43 plaque size was reduced by 2.2-fold (from 1 ± 0.08 to 0.45 ± 0.2) in chagasic patients, this being the main characteristic of the observed disassembly of this group (Fig. 4B), thus corroborating the observed loss of Cx43 plaques (Fig. 3D, G). To compare the intensity of cardiac fibrosis and positive TGF-β-reactive cells with Cx43 expression, we analysed in human heart sections the presence of fibronectin and PS2, markers of fibrosis and of active TGF-β signalling, respectively. When compared to healthy controls (Fig. 3B, C), higher levels of pericellular fibronectin staining (Fig. 3H) and nuclear staining to PS2 (Fig. 3I) were observed on the sections of cardiac tissue from chagasic patients in parallel to the reduced Cx43 staining (Figs 3G, 4A, B). The main clinical data and quantitative analysis of Cx43 size and expression from each patient are shown in Table II. The histology analysis and immunolabelling of T. cruzi antigens and of inflammatory infiltrates of these patients have already been studied and described elsewhere (Higuchi et al. 1997, 1999, Reis et al. 1997, Palomino et al. 2000). The normal pattern of Cx43 staining in human ventricular myocardium (healthy donors) has already been determined (Jiang et al. 2005), thus only one heart was used as a control due to ethical constraints. Heart weight from the chagasic group was higher than the non-infected controls (mean weight, 457.5 g compared to 235 g, respectively). Data in Table II allowed a correlation study between Cx43 protein expression levels and heart weight (Fig. 4C), depicting a clear and significant negative correlation between these parameters (r2 = 0.8244). Fewer Cx43 plaques were observed in heavier hearts. We could not predict a correlation with the presence of cardiac insufficiency and/or myocarditis with the number of Cx43 plaques because all chagasic patients presented with myocarditis; however, the Cx43 level was not homogeneous (Table II). DISCUSSION In the heart, gap junctions mediate electronic current flow, thereby coordinating the spread of excitation and subsequent contraction throughout the myocardium (Spray & Burt 1990). Intercalated discs join neighbouring cardiomyocytes and provide mechanical adhesion as well as electrical and metabolic coupling. Protein changes in these junctions, particularly in gap junctions, are known to be involved in pathological features (Severs 2001). The disturbance of gap junction signalling could lead to altered impulse propagation between cardiomyocytes and to ventricular arrhythmogenesis in the myopathic heart (Peters et al. 1993). In CCC, one of the main complications is heart failure in association with extensive fibrosis (Higuchi et al. 1999), high circulating TGF-β levels (Araújo-Jorge et al. 2008) and arrhythmia (Casado et al. 1990). To examine the effect of TGF-β on Cx43 protein expression, we employed an in vitro model of cardiomyocytes submitted to treatment with the cytokine, its inhibitor, or T. cruzi infection. Our findings demonstrated that TGF-β addition altered the Cx43 protein expression in non-infected myocytes, resulting in reduced organisation of gap junctions, which led to a punctate, diffuse and non-uniform pattern of staining. We observed a decrease in Cx43 staining in infected cells and their neighbours, thus confirming what has already been demonstrated (Campos de Carvalho et al. 1992, Adesse et al. 2008). These results predicted that direct T. cruzi infection of heart cells could slow cardiac conduction and alter the pattern of impulse propagation as a consequence of decreased strength of electrotonic coupling. Increased local production and secretion of TGF-β-1, as we demonstrated, could also contribute to impaired gap junctional communication in cardiomyocytes. The involvement of TGF-β signalling in the mechanism of Cx43 decrease was verified when non-infected cardiomyocytes also developed a punctate Cx43 labelling after exposure to exogenous TGF-β, similar to the pattern observed in infected cells. It was further reinforced by the interesting results obtained with the ALK5 inhibitor, SB-431542, which completely reversed the effect of TGF-β and of T. cruzi infection on Cx43 expression as detected by immunoreaction and by western blot analysis. These results indicate a possible role of endogenously secreted TGF-β as an autocrine/paracrine agent. This result suggests that TGF-β produced in infected cultures could affect both infected and non-infected cells and induce the diffuse pattern of Cx43 staining and could be implicated in the decrease of Cx43 protein expression in infected cells. In addition, this decrease may be related to a general instability of global cytoplasmic mRNA that is commonly associated with T. cruzi intracellular proliferation (Pereira et al. 2000). In macrophages, it has been demonstrated that the altered electrical conduction between cells was due to a different pattern of Cx43 localisation rather than altered Cx43 expression levels (Fortes et al. 2004). Our present data agree well with data obtained in chagasic patients, who exhibited severe diffuse fibrosis (increased fibronectin expression associated with a strong nuclear stain for PS2), indicating higher levels of active TGF-β (Araújo-Jorge et al. 2008). In this study, we observed a significant down-regulation of Cx43 protein expression, a reduction in the length of Cx43 plaques and an altered distribution of these plaques in the chagasic group. These observations may account for the abnormal impulse conduction commonly observed in electrocardiogram profiles of Chagas disease patients and for myocytolisis leading to intense heart fibrosis contributing to cardiomegaly, as shown in Fig. 4C. These data are consistent with previous studies of patients with heart failure of different origins (Kostin et al. 2003). Disruption of the normal pattern of gap junction distribution also occurs in myocytes bordering areas of fibrotic myocardial damage and in areas of myocardial inflammation, where impairment of the impulse conduction was observed (Spach & Boineau 1997). T. cruzi infection is a strong inducer of TGF-β expression in several cell types (Waghabi et al. 2002). Higher amounts of TGF-β could be implicated either in the gap junction disassembly or in the observed extensive fibrosis, but probably plays a role in both phenomena. Higher levels of TGF-β and disorganised expression of Cx43 could both act in synergy to promote heart fibrosis, thus leading to a more severe cardiopathy. In summary, our results indicate that an important component of the autocrine myocyte response to TGF-β secretion is the change of Cx43 protein expression, which contributes to the disturbance of cardiac electrical conduction. Taken together, the results shown here introduce more elements to the complex picture of heart fibrogenesis and arrhythmia, thus connecting TGF-β and Cx43 dysregulation as key events in Chagas disease cardiomyopathy. ACKNOWLEDGEMENTS To Mr. Daniel Ciantar and Dr. Carlos Alberto Bizzarro Rodrigues, for the confocal analysis, to Dr. Gillian E. Knight, for editorial assistance, and Dr. Joseli Lannes-Vieira, for her generous gift of anti-Cx43 antibodies. REFERENCES
Copyright © 2009 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc09222f2.jpg] [oc09222t2.jpg] [oc09222f3.jpg] [oc09222f1.jpg] [oc09222t1.jpg] [oc09222f4.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}