 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
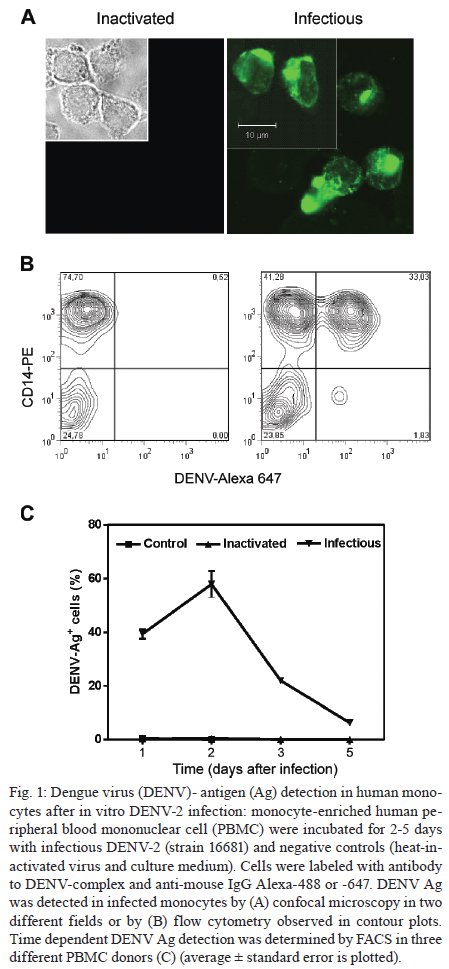
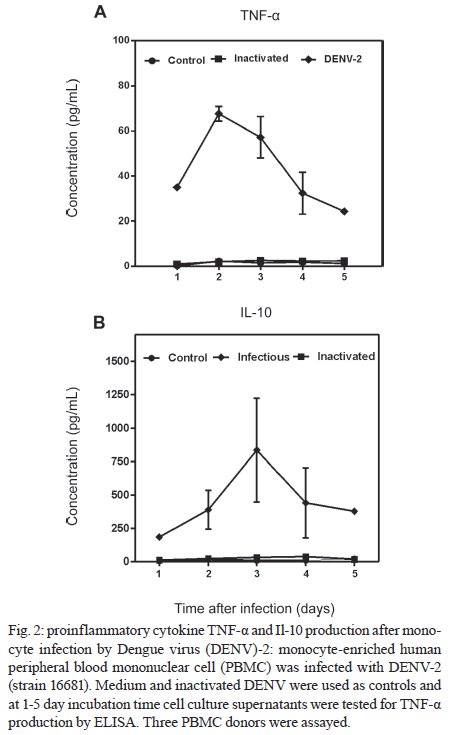
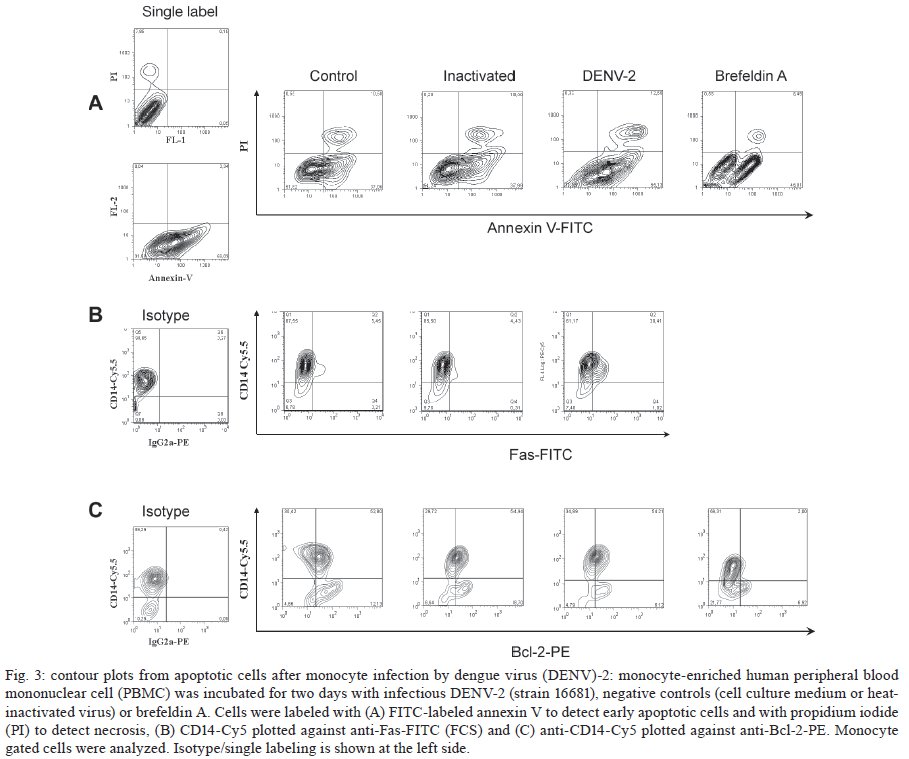
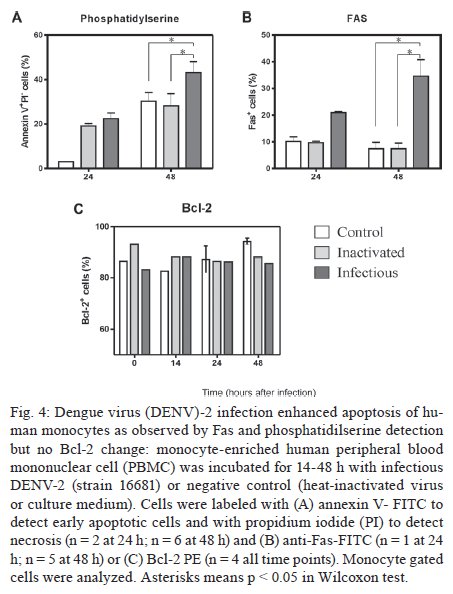
Memórias do Instituto Oswaldo Cruz, Vol. 104, No. 8, 2009, pp. 1091-1099 ARTICLES Dengue-2 infection and the induction of apoptosis in human primary monocytes Amanda Torrentes-Carvalho; Elzinandes L Azeredo; Sonia RI Reis; Alessandro S Miranda; Mariana Gandini; Luciana S Barbosa; Claire F Kubelka+ Laboratório
de Imunologia Viral, Instituto Oswaldo Cruz-Fiocruz, Av. Brasil 4365, 21045-900
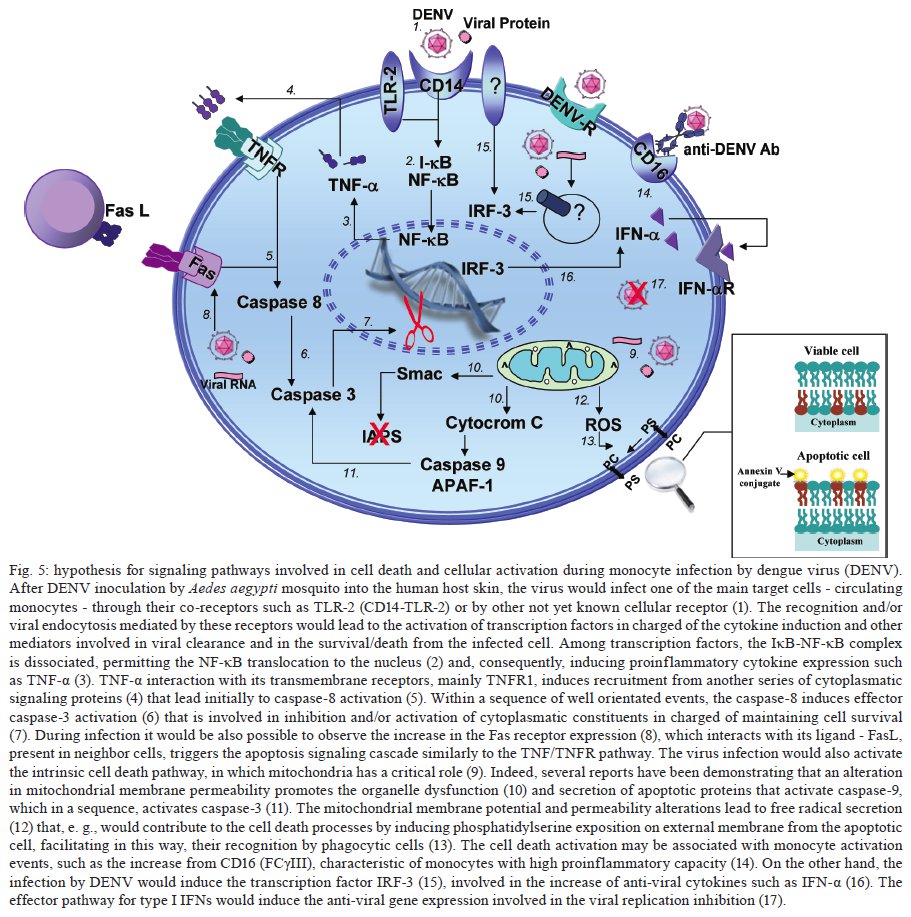
Rio de Janeiro, RJ, Brasil Financial support: IOC-FIOCRUZ, DECICT/ CNPq, FAPERJ, ICGEB Received 28 June 2009 Code Number: oc09223 ABSTRACT Monocytes/macrophages are important targets for dengue virus (DENV) replication; they induce inflammatory mediators and are sources of viral dissemination in the initial phase of the disease. Apoptosis is an active process of cellular destruction genetically regulated, in which a complex enzymatic pathway is activated and may be trigged by many viral infections. Since the mechanisms of apoptotic induction in DENV-infected target cells are not yet defined, we investigated the virus-cell interaction using a model of primary human monocyte infection with DENV-2 with the aim of identifying apoptotic markers. Cultures analyzed by flow cytometry and confocal microscopy yielded DENV antigen positive cells with rates that peaked at the second day post infection (p.i.), decayed afterwards and produced the apoptosis-related cytokines TNF-α and IL-10. Phosphatidylserine, an early marker for apoptosis, was increased at the cell surface and the Fas death receptor was upregulated at the second day p.i. at significantly higher rates in DENV infected cell cultures than controls. However, no detectable changes were observed in the expression of the anti-apoptotic protein Bcl-2 in infected cultures. Our data support virus modulation of extrinsic apoptotic factors in the in vitro model of human monocyte DENV-2 infection. DENV may be interfering in activation and death mechanisms by inducing apoptosis in target cells. Key words: dengue - apoptosis - monocytes - human The dengue fever emergency has become one of the most dramatic public health issues in Latin America. Epidemics are presenting increased severity and frequency and are continuously expanding geographically (Gubler 2002). In Brazil, the disease is endemic and during the last five years the region has accounted for 70% reported cases in the Americas (Nogueira et al. 2007). Infection with the Dengue virus (DENV) causes a disease spectrum ranging from asymptomatic to shock and hemorrhages, with a risk of lethality. Acute vascular permeability may be followed by abnormal vascular homeostasis. Clinical manifestations include plasmatic leakage, bleeding tendency and liver commitment followed by mild seric transaminase elevation. The physiopathology of the disease may be explained as result of infected monocyte activation by the production of factors that act to increase the vascular permeability and disturb the coagulation system. Pleural effusion, ascites and hemoconcentration are signs of intravascular volume loss. Such factors promote rapid progression to shock (Gibbons & Vaughn 2002). Mononuclear phagocytes are considered the main targets for viral replication (Jessie et al. 2004, Neves-Souza et al. 2005). Cells such as B lymphocytes, monocytes, hepatocytes and dendritic cells are described as potential virus targets and may undergo programmed cell death when infected in culture. Reports show apoptotic induction in DENV infected endothelial cells elevates IL-8 and RANTES production, which are chemokines related to inflammatory processes during cell migration (Avirutnan et al. 1998). Monocytes/macrophages and dendritic cells are responsible for the dissemination of the virus after its entrance into the dermis by mosquito bite (Kou et al. 2008). Monocytes and T lymphocytes are activated during human infection, producing several pro-inflammatory mediators and may have amplified activation during secondary infection to become sources of intense cytokine production. Several investigations have followed patients during acute infection to monitor their plasma cytokine content and found that TNF-α is among the first detected cytokines in patients as well as in peripheral blood mononuclear cell (PBMC) cultures (Bethell et al. 1998, Braga et al. 2001, Reis et al. 2007). In general, these mediators may promote platelet and endothelial activation, culminating in thrombocytopenia, vascular permeability, hypotension and shock, as observed in severe cases of dengue (Fink et al. 2006). The TNF superfamily cytokines mediate their effects by activating transcription factors, including NF-κB, which culminate in apoptotic processes and/or cellular proliferation (Karin & Lin 2002). Programmed cell death is an essential event during normal life in addition to being involved in pathophysiological processes that trigger disease (Lemasters 2005). Apoptosis consists of a controlled process of cell death that is orchestrated by a cascade of biochemical events, leading to morphologic changes and eventually to cell death (Bruchhaus et al. 2007). It is a physiologic mechanism that may exert pathogenic effects as well. Cells with damaged DNA can be removed without harm via the activation of a complex enzymatic chain of intracellular signaling systems that lead to the destruction of components that are essential to cell survival. These events occur in response to a variety of signals and stimuli, both internal and external, including growth factor deficiency, hormones, cytokines and suicidal cells that activate death receptors and endogenous proteases (Lemasters 2005). During the apoptotic process, changes in the cellular membrane expose phosphatidylserine to the external cell surface; subsequently, there is cleavage of the DNA double helix by endogenous endonucleases, among other changes (Vermes et al. 1995, Merchant et al. 2001). Apoptosis is usually induced by an extrinsic signal transduction triggered by a death receptor such as TNF receptor (TNF-RI), TRAIL-RI/II and/or Fas (CD95/APO-1) binding to TNF-α, TRAIL or Fas ligand. After signaling, an enzymatic cascade leads to the activation of a series of cysteinyl aspartate proteases known as caspases (Feig & Peter 2007) and then to cell degredation. However, the apoptotic process can be triggered by endogenous events as well. It is well-known that mitochondria play a central role in mediating intrinsic apoptotic signals. Changes in the external membrane mitochondrial potential and the production of reactive oxygen species are related to initial apoptotic events. When death signals reach the mitochondria, they lead to a collapse in the membrane potential that results in organelle rupture and release of pro-apoptotic proteins to cytoplasm, inducing caspase activation (Loeffler & Kroemer 2000, Gupta 2003). The mitochondrial apoptotic program is also regulated by several Bcl-2 family members that are known to have an active role in inducting/inhibiting cell death (Gross 2001, Dejean et al. 2006). DENV may be inducing apoptosis and disseminating its viral progenies to neighbor cells through phagocytized apoptotic bodies; alternatively, it may be acting to inhibit cell death in order to ensure virus survival and replication. Considering that phagocytosis of apoptotic cells is essential in preventing a deregulated inflammatory process and inducing specific immunological responses, the induction of apoptosis may be an attempt by the host immune system to limit the extent of the infection. Since monocytes/macrophages represent the main targets for infection by DENV and have an essential role in developing disease severity, it would be extremely useful to reveal processes that modulate the virus-cell interaction effect, with an emphasis on cell death mechanisms. To that end, we here studied an in vitro infection model in which primary human monocytes are infected with DENV-2, produce TNF-α, IL-10 and are expressing apoptotic markers such as phosphatidylserine exposition and Fas upregulation, indicating that apoptotic processes are occurring. MATERIALS AND METHODS Virus strain, cell cultures, virus stock preparation and titration - DENV, serotype 2, strain 16681 was provided by Dr. SB Halstead (Naval Medical Research Center, USA). For virus stock preparation, an Aedes albopictus C6/36 cell clone was grown as a monolayer culture, as previously described by Reis et al. (2007). Virus stock was titrated by serial dilution cultures in microtiter plates and detected by immunofluorescence, as previously described (Miagostovich et al. 1993). Virus titer was calculated as 50% tissue culture infectious dose (TCID50/mL). The inactivated virus was prepared by incubating the inoculum for 30 min/56°C. Titrated virus stock was used in culture at a final concentration of 1.37 x 108 TCID50/mL. In vitro purification of PBMC - Human peripheral blood was obtained from the Hospital Universitário Clementino Fraga, Rio de Janeiro. PBMCs were isolated from healthy donors' buffy coats by centrifugation on a density gradient (350 g/30 min in Ficoll-Paque Plus, Amersham Biosciences Corp) according to standard procedures. Cell suspensions (1 x 106 cells/mL) in RPMI 1640 medium containing 200 mM glutamine, 100 U/mL penicillin and 10 mg/mL streptomycin, were incubated at 37°C under humid atmosphere with 5% CO2 and allowed to adhere on uncoated polystyrene flasks (150 cm3) during 90 min for monocyte enrichment. Non-adherent cells were gently removed by washing after which the adherent cells were detached by mechanical cell harvesting with cell scrapers in cold cell culture medium. Cell viability was verified in culture by Trypan blue exclusion and presented > 95% viability. Enriched monocytes were suspended in RPMI 1640 medium supplemented with 10% FCS and seeded at 1 x 106 cells/mL on 24-well plates or 16-well cell culture chambers (Labtech, Nunc). Infection of adherent PBMCs - After an overnight incubation, cell cultures were infected using a DENV-2 diluted inoculum (300 μL), inactivated virus or maintained with culture medium. After 2 h incubation for adsorption, culture supernatants were replaced with medium containing 2% FCS and further incubated for 24-120 h. Wells were set in triplicates for each different parameter in culture. After 48 h, supernatants were collected and stocked at -20ºC until cytokine measurement and cells were recovered for viral antigen (Ag), co-stimulatory, intrinsic and extrinsic apoptotic marker determination by flow cytometry. Determination of viral Ag and specific co-stimulatory marker by flow cytometry in human monocytes - Cells were recovered by scratching with a plastic microtip using cold medium and were set at 1 x 106cells/microtube; they were then centrifuged (350 g, 5 min) and washed once with 1 mL PBS pH 7.4 with 2% FCS and 0,01% NaN3. Single or double labeling of infected and uninfected adherent PBMCs was performed. For surface labeling, cells were blocked with 5% inactivated plasma in PBS/BSA at 4ºC/30 min and incubated with FITC or Cy5.5-labelled antibodies against CD14 (Southern Biotheck), a specific molecule for monocyte population. Matching isotype antibodies were used as a negative control. Intracellular staining after infection was performed as previously described (Neves-Souza et al. 2005), with slight modifications. Briefly, cells were fixed with 0.5 mL cold paraformaldehyde at 2% in PBS for 10 min and, after centrifugation, membrane permeabilization was carried out with 1 mL 0.1% saponine in PBS with FCS and NaN3. Cell surface unspecific receptors were blocked with 5% inactivated human plasma in PBS/BSA at 4ºC/30 min. Primary monoclonal antibodies reactive to Dengue Complex (Chemicon) or isotype anti-mouse IgG2a diluted in PBS with saponine, FCS and NaN3 were added to cells for a 60 min incubation. Cells were washed once with 1 mL PBS containing FCS and NaN3 and further incubated with a secondary anti-mouse IgG1 labeled with Alexa 488 (BD Pharmingen) for 30 min. Alternatively, for double staining with CD14 and DENV, cells were first surface labeled with PE-labeled antibodies to CD14 (DAKO) and then intracellularly labeled with monoclonal antibody to Dengue Complex-Alexa 647 (coupled with Molecular Probes labeling kit). Cells were then acquired (5,000 events for gated monocytes) on a FACSCalibur flow cytometer (Beckon & Dickinson). Analysis was performed using FlowJo Software (TreeStar Inc). Assessment of apoptosis - Apoptotic detection assays were carried out by surface labeling with the Ca2+ dependent phosphatidylserine-binding protein annexin V. Uninfected and infected cells were recovered by scratching with plastic microtips using cold medium and were set at 1 x 106/microtube; they were then centrifuged (350 g, 5 min) and washed once with PBS 1X. Cells were labeled by incubation with Annexin V-FITC and propidium iodide (PI) in a binding buffer (Apoptosis detection Kit I # 556547, BD Pharmingen) for 15 min at RT in the dark, according to the manufacturer's instruction. For extrinsic (surface) apoptotic molecules labeling, cells were blocked and incubated with FITC-labeled antibodies to Fas (Southern Bioteck), a death receptor. Intracellular staining was performed for detection of Bcl-2, an intrinsic anti-apoptotic protein, using PE-labeled antibodies to Bcl-2 (Caltag) as described above. Specific isotypes were used as negative controls. For an apoptotic positive control, culture wells were treated with Brefeldin A (Sigma), a fungal macrocyclic lactone that is a potent inhibitor of membrane trafficking and vesicular transport and which therefore induces apoptosis when incubated with cultures at low concentrations for long time periods (Shao et al. 1996, Guo et al. 1998). Stained cells were acquired (5,000 for gated monocytes) on a FACSCalibur flow cytometer. Analysis was performed using FlowJo Software. Confocal microscopy - Cells were isolated and cultured in Labtech microchambers (Nunc). Two days after infection, as described above, supernatants were recovered and adherent cells were washed once with PBS 1X and fixed with 0.05 mL cold paraformaldehyde at 2% and Triton 0.2% (Sigma) at 4ºC/30 min. After fixation, wells were washed with PBS/BSA and blocked with 5% inactivated plasma in PBS/saponine at 4ºC/30 min. Subsequently, cells were incubated with Dengue Complex-reactive primary monoclonal antibodies or isotype anti-mouse IgG2a (Chemicon) for 37ºC/60 min in a humid chamber. Cells were washed twice with PBS/BSA and further labelled with a secondary anti-mouse IgG1 coupled to Alexa 488 (BD Pharmingen) for 37ºC/60 min in a humid chamber. The microchambers were washed with distillated water and the chambers were removed from slides. Slides were dried and mounted in glycerol with 2% anti-fading agent. Cells were observed under a fluorescence microscope LSM510-META (Zeiss) using excitation/emission (488 nm/ 540 nm - green fluorescence dye) for DENV. Digital pictures of nuclei and virus were obtained for each stain using a CCD camera. Ethics - Procedures performed during this work were approved by the Ethical Committee of the Fundação Oswaldo Cruz, Brazilian Health Ministry (recognized by the Brazilian National Ethical Committee) (111/00). Statistical analyses - Data were first tested for normality with the GraphPad Prism version 5 for Windows GraphPad Software (http://www.graphpad.com) in order to determine the significance of differences in dengue Ag, Annexin V and Fas death receptor positive cell numbers under several culture treatment conditions. Data values passed the Kolmogorow-Smirnov normality test and were evaluated for significance using the one-way ANOVA (more than two groups). Altered parameters were considered significant at p < 0.05. RESULTS DENV Ags are expressed after monocyte infection; proinflammatory cytokine TNF-a production peaked at two days and IL-10 at three days post infection (p.i.) - Monocytes infected with DENV-2 (strain 16681) were cultured for five days in order to evaluate cell infection and cytokine production. Both DENV intracellular Ag and secreted cytokines present in the cell culture supernatant were determined daily. Cytoplasmatic virus localization was observed by confocal microscopy with immunofluorescent labeling (Fig. 1A). Infected cells are CD14+ monocytes as represented by contour plots during flow cytometry analysis within the monocyte gate (Fig. 1B). Increased DENV Ag+ cell rates were detected by flow cytometry until the second day p.i., when peak values were reached. After this time point, DENV Ag+ cell percentages decreased (Fig. 1C). Monocytes from 12 PBMC healthy donors showed mean virus infection percentage of 40 ± 4% on the second day p.i. with a minimum of 18% and a maximum of 58% DENV Ag+ cells. TNF-α is a pro-inflammatory cytokine that followed the same kinetic pattern as cell infection, with maximum levels at day two and a subsequent gradual decay from 3-5 days after infection (Fig. 2). IL-10 is an anti-inflammatory cytokine that may be induced by TNF-α as a regulatory feedback mechanism and reached its peak a day later. Both are detected in cell culture supernatants and are most likely produced as a result of cell activation by the virus and stay in culture longer than viral Ags detected intracellularly. Most experiments performed showed that intracellular virus was already high by the first day, peaked at the second day and then decayed abruptly and from the third day onwards was either undetected or detected at low rates. In contrast, cytokines displayed statistically significant differences in their levels for 3-5 days. Changes related to programed cell death in monocytes infected with DENV-2 Membrane changes exposing phosphatidylserine occur on infected monocytes - In order to evaluate if DENV was inducing apoptosis in human monocytes, we searched for an early marker of cell death, the display of phosphatidylserine at the cell membrane, by labeling with annexin V and PI. The presence of annexin V without PI labeling is used to identify apoptotic and autophagic cells at early stages of cell death, when annexin V binding is exclusive. Double-labeling occurs at later stages of apoptosis and in subsequent necrotic processes. Brefeldin A, a lactone antibiotic produced by fungal organisms that induces apoptosis, was added to cell cultures as positive control. Two days after infection of monocytes with DENV-2, cells were recovered for flow cytometry analysis. Fig. 3A shows contour plots representing annexin V and PI labeling on monocytes incubated with cell culture medium, inactivated virus, infectious DENV-2 or brefeldin A. Infected cells presented an increase in single-labeled annexin V+ cells as compared to cultures incubated with inactivated virus or medium. No significant differences were observed in the frequency of cells that underwent apoptosis followed by necrosis (annexin-V+ PI+) when inactivated was compared to infectious virus. Cells incubated only with inactivated virus displayed equivalent percentages as those incubated with medium, indicating that physiological culture conditions are permitting cells to die spontaneously or recovery procedures may damage some cells. Nevertheless, the virus enhanced the apoptotic process that was occurring in culture. Data from five different PBMC donors for annexin V+PI- cells are plotted in Fig. 4A, showing the significant difference between cultures exposed to infectious DENV-2 and controls. Differences in early apoptotic markers between infected and uninfected cells were not significant until 48 h. Fas receptor expression is detected on infected monocytes - The Fas cell surface receptor (CD95) can be stimulated by agonists or the natural ligand FasL, recruiting proteins with distinct functions such as cytosolic adaptor proteins that ultimately bind to the initiating caspase (caspase 8) and trigger apoptosis (Limjindaporn et al. 2007). DENV-2 infected monocytes were also analyzed in order to detect the Fas expression on cell surface. By the first day after infection, Fas+ cells were already more common in DENV-2 exposed cells than in controls. At two days after infection, Fas+ cell frequencies were significantly increased in the presence of the infectious virus, as detected in six different PBMC donors (Fig. 4B). Bcl-2 oncoprotein expression is not altered in infected monocytes - The proto-oncogene Bcl-2 works as an important repressor of programmed cell death. We investigated whether DENV-2 infected monocytes, apparently more susceptible to death, might have altered Bcl-2 expression and therefore an altered degree of inhibition of apoptosis. We detected Bcl-2 by labeling monocytes from 1-5 days after DENV-2 infection and analysed the cells by flow cytometry (data not shown). Bcl-2 expression remained unchanged during this period, although alternative treatment with brefeldin A was able to downregulate this protein, as can be observed in Fig. 3C (showing data from the first 48 h). In order to ascertain whether cells were infected or not with DENV, we performed double-labeling with antibodies directed against Bcl-2 and DENV. We detected double labeling in 39 ± 4% cells at 24 h and 50 ± 2% at 48 h, in contrast to negative double label in controls (< 1%), indicating that the presence of the DENV-2 does not interfere with Bcl-2 expression; most infected cells are still expressing this protein and the apoptosic mechanisms appear to act independently from Bcl-2 regulation. DISCUSSION Mononuclear phagocytes such as monocytes and dendritic cells are the main DENV target cells, both in vitro and in vivo and their interaction with the virus has been shown to be relevant to the host response to infection (Ha-lstead & O'Rourke 1977, Wu et al. 2000, Neves-Souza et al. 2005). After signaling pathway activation by viruses, these cells have upregulated surface activation molecules and begin producing cytokines and chemokines. Modulation of the innate and adaptative immunity interface occurs as a result (Frankenberger et al. 1996, Bosch et al. 2002, Strauss-Ayali et al. 2007), likely influencing the protection and/or pathology onset during infection. DENV replicates in human primary peripheral monocytes (Halstead & O'Rourke 1977, Chen & Wang 2002). We have previously infected human monocyte cultures with DENV-2 Asian strain 16681 (Sydow et al. 2000) and other DENV strains (Neves-Souza et al. 2005). Here we show a time-dependent detection of DENV Ags starting as early as the first day after infection, with a detection peak at 48 h and a subsequent decay. These results resemble other reports using this virus strain in dendritic cells, although peak virus production may be related not only to the strain type but also with the inoculum concentration (Cologna & Rico-Hesse 2003). Intracellular virus detection was related to TNF-α extracellular production, with a maximum production at the second day. TNF-α is probably produced as a result of virus replication and remains in culture, being detected at significant levels at five days, by which point intracellular virus is found at low levels. It is likely that non-infected monocytes are activated and become important targets for TNF-α production at the later stage of infection. Programmed cell death is an essential regulatory process in the immunological system. During immune responses to viral infections, apoptosis may occur as a direct pathogenic mechanism of viral escape, allowing apoptotic bodies containing virus to disseminate infection to neighbor cells. Alternatively, apoptosis may be used as an immune mechanism to induce opposed effects, becoming deleterious to viral progenies and representing an appropriate response from host to limit viral replication (Servet-Delprat et al. 2000). Apoptosis can be either Ag-dependent, initiated by death receptor signaling and/or activating intracellular pathways, or it may occur by negligence when Ag-specific receptors are not stimulated. When the cellular microenvironment is depleted of survival factors, another type of programmed cell death occurs, a phenomenon known as authophagy (Lemasters 2005, Feig & Peter 2007). Moreover, modulation of programmed cell death plays an important role in several pathologies related to viral infections such as AIDS and measles (Oyaizu & Pahwa 1995, Servet-Delprat et al. 2000). DENV replication seems to be essential to induce both phagocytosis and apoptosis in human monocytes, as reported by Espina et al. (2003). However, these authors showed no quantitative evidence about apoptotic events. Considering that monocytes are among the main target cells for DENV, we investigated markers of programmed cell death during human primary monocyte infection, as well as the production of related cytokines. During phagocytosis, the virus-cell interaction may lead to production of cytoplasmatic lysosomal components that can result in deleterious effects for both the virus and host cells. Labeling cells with annexin V, which binds to the exposed phosphatidylserine found on apoptotic cells and the simultaneous exclusion with propidium iodine evaluated by flow cytometry allowed us to quantify early apoptosis after two days of DENV infection in monocytes. This observation matches with the peak production of TNF-α, which is a proinflammatory cytokine that plays an important role in the induction of apoptosis. We suggest that the maturation process and the cytokine production by these cells are activation pathways involved in apoptotic processes. We also observed that some monocytes cultured without infectious virus die, although at lower rates. It is possible that they are undergoing cell death processes by mechanisms such as authophagy if survival factors are not present at sufficient concentrations in cultures (Yu et al. 2006). Among TNF family receptor series, the Fas receptor is related to the pathenogenesis of viral infection such as measles and hepatitis B (Servet-Delprat et al. 2000, Wang et al. 2004). Dendritic cells infected by measles virus present upregulated expression of Fas, becoming more susceptible to apoptosis mediated by the Fas/FasL system. Also, a protein from the DENV capsid induces apoptosis in HepG2 cells in a Fas-dependent fashion (Limjindaporn et al. 2007). Fas signaling induces IL-8, TNF-α and IL-1β production in mononuclear phagocytes, modulating pro-inflammatory responses by a cas-pase-dependent pathway (Park et al. 2003). IL-10 is a potent anti-inflammatory cytokine. This factor triggers apoptosis in monocytes via CD95 system and monocytes undergoing spontaneous apoptosis in vitro become a potent source of IL-10 (Schmidt et al. 2000, Bzowska et al. 2002). Elevated Il-10 levels have been detected in patients with dengue fever and often have been found to be associated with increased severity (Green et al. 1999, Azeredo et al. 2001). We demonstrated here for the first time that human monocytes infected in vitro by DENV have upregulated Fas expression concomitant with the viral peak, indicating that programmed cell death induced by DENV in our study model may be influenced by this extrinsic apoptotic pathway. It is possible that this process leads to extracellular release of virus, facilitating neighbor cell infection, in addition to inducing simultaneously monocyte maturation. Fas and DENV Ag double cell labeling in DENV-2 infected cultures shows that Fas expression may occur in DENV-Ag positive and negative cells. DENV-Ag is detected in Fas-negative cells (data not shown). Although our data indicate that apoptosis is induced in monocytes during Fas expression, we cannot exclude the possibility that TNF-α/TNFR receptor, cytokines and/or other mechanisms are participating in apoptotic processes mediated by DENV. Azeredo at al. (2001) reported elevated circulating TNF-α and TNF soluble receptors in patients with acute dengue fever. The cytoplasmatic expression of the proto-oncogene Bcl-2 plays a role in increasing cell survival and has contrary effects to Fas during apoptosis (Boudet et al. 1996, Mo-lica et al. 1996). However, no significant changes were detected with regard to Bcl-2 expression in monocytes infected with DENV-2, in contrast to what we observed in CD8 positive T-lymphocytes from dengue patients, which presented low levels of Bcl-2 (Azeredo et al. 2006). The present data suggest that other anti-apoptotic proteins, such as Bcl-xL, might be negatively regulated during infection in monocytes. According to our data and other relevant studies, we present here a model for DENV infection in monocytes in which complex interactions between the virus and host cell occur, leading to the activation of transcription factors, cytokines and enzymatic factors. These interactions may induce not only pathologic proinflammatory responses that influence the severity and progression of the disease, but also virus clearance and apoptotic pathways that could be controlling infection by anti-viral mechanisms [(Chen et al. 1999, Diamond et al. 2000, Bielefeldt-Ohmann et al. 2001, Grandvaux et al. 2002, Navarro-Sanchez et al. 2003, Neves-Souza et al. 2005, Azeredo et al. 2006, 2009, Seth et al. 2006, Chareonsirisuthigul et al. 2007, Reis et al. 2007, Durbin et al. 2008, Kou et al. 2008 (see details in Fig. 5)]. The present study may, therefore, contribute to a better understanding of the cell activation mechanisms triggered during DENV infections occurring in human target cells. Further investigations will be required in order to define in detail the apoptotic pathways involved in dengue fever and to determine whether or not there is a potential benefic role from apoptosis in the resolution of infection. ACKNOWLEDGMENTS To Bernardo M Pascarelli, by helping in confocal image analysis and uptake at the Confocal Microscopy Technological Platform, Fiocruz, and to Luzia Maria de Oliveira Pinto, for suggestions and encouragement. Amanda Torrentes de Carvalho was a fellowship from CNPq. REFERENCES
Copyright © 2009 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc09223f4.jpg] [oc09223f5.jpg] [oc09223f2.jpg] [oc09223f3.jpg] [oc09223f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}