 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
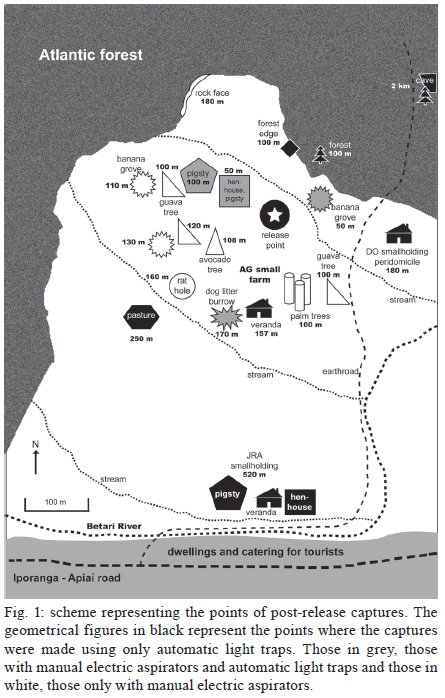
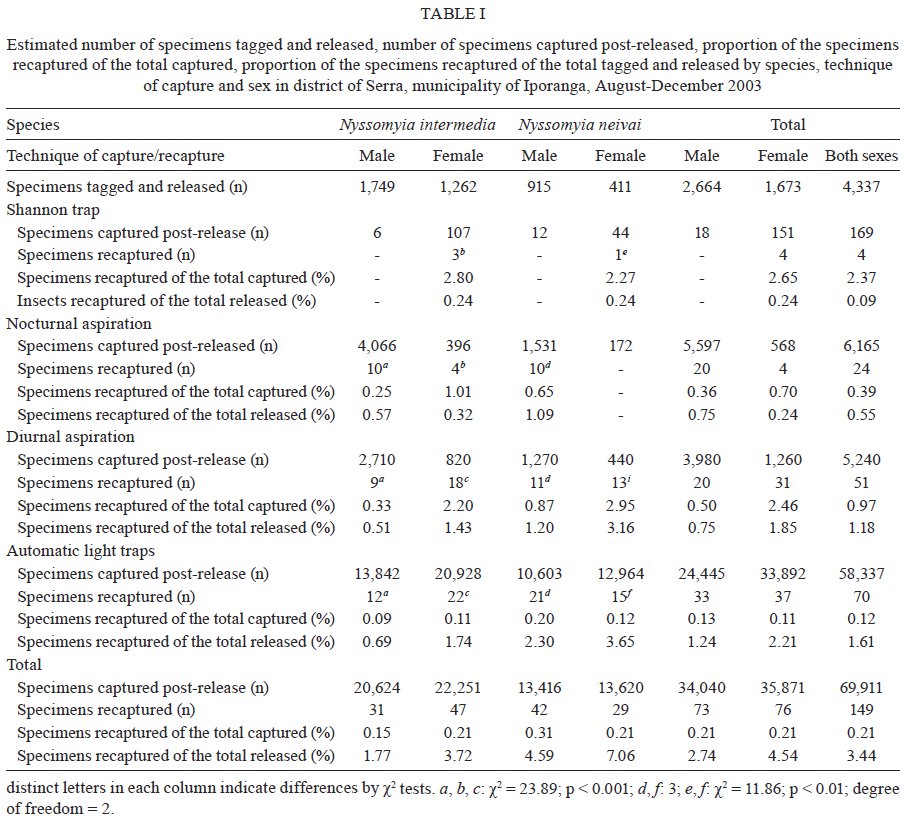
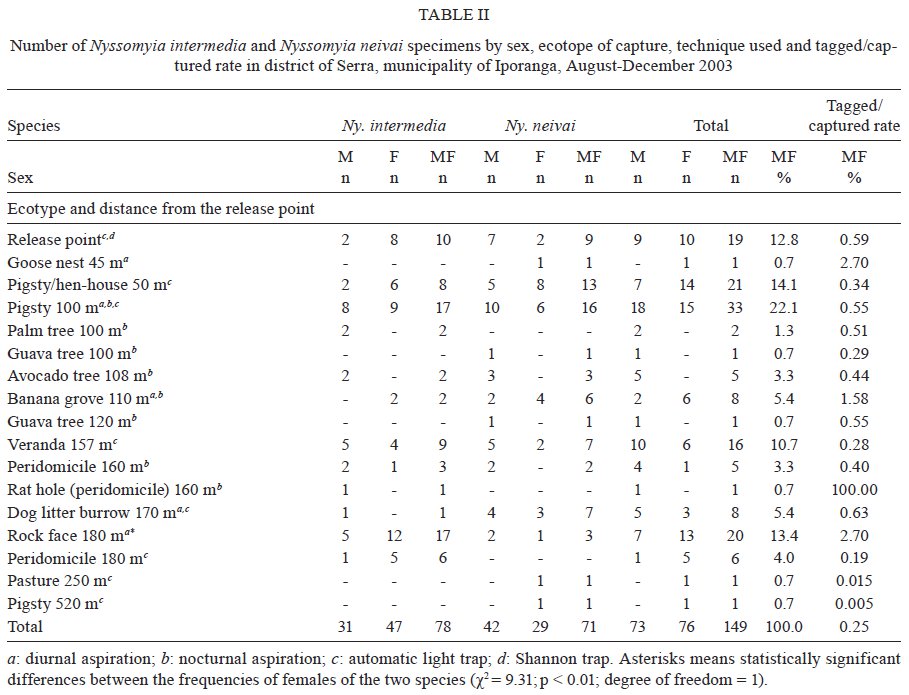
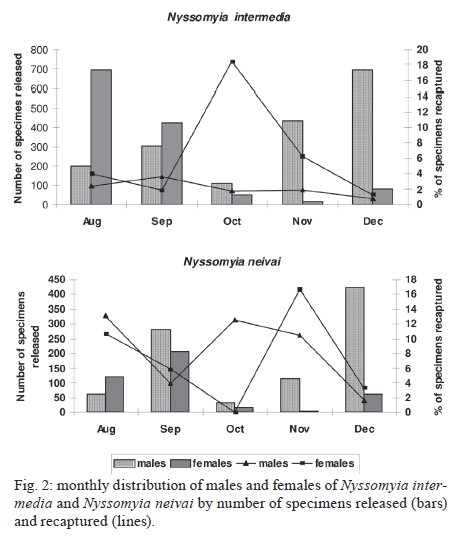
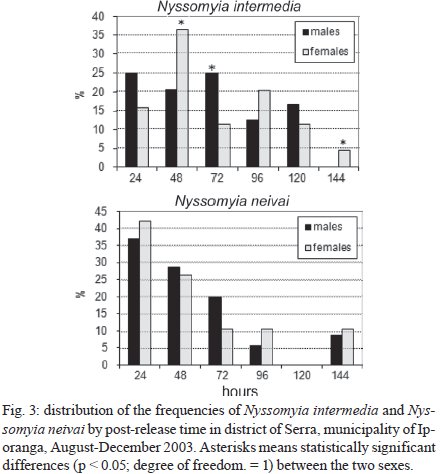
Memórias do Instituto Oswaldo Cruz, Vol. 104, No. 8, 2009, pp. 1148-1158 ARTICLES Dispersal and survival of Nyssomyia intermedia and Nyssomyia neivai (Diptera: Psychodidae: Phlebotominae) in a cutaneous leishmaniasis endemic area of the speleological province of the Ribeira Valley, state of São Paulo, Brazil Eunice AB GalatiI, +; Mauro Breviglieri FonsecaI; Ana Maria MarassáI, II; Edna Fátima M BuenoI IDepartamento
de Epidemiologia, Faculdade de Saúde Pública, Universidade de
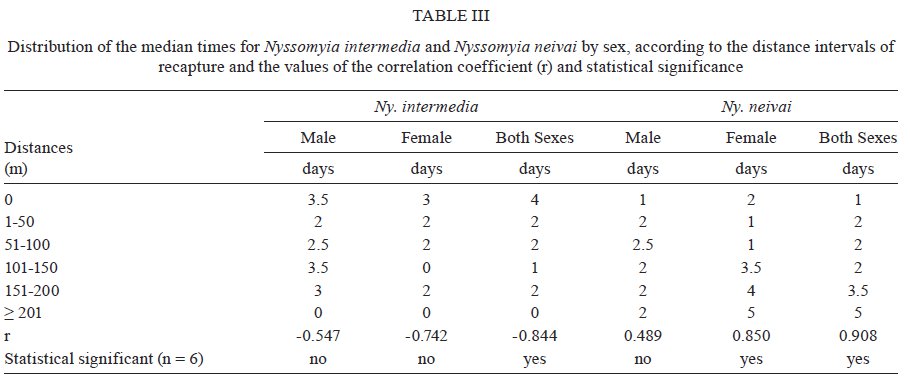
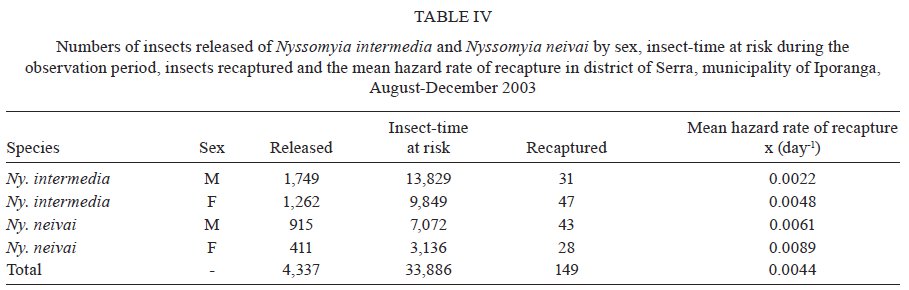
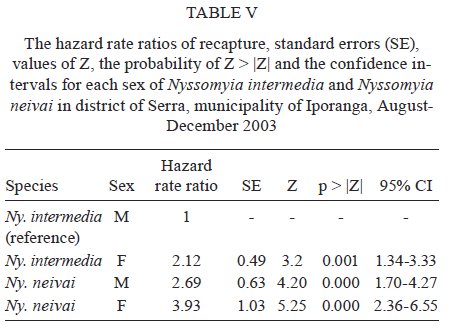
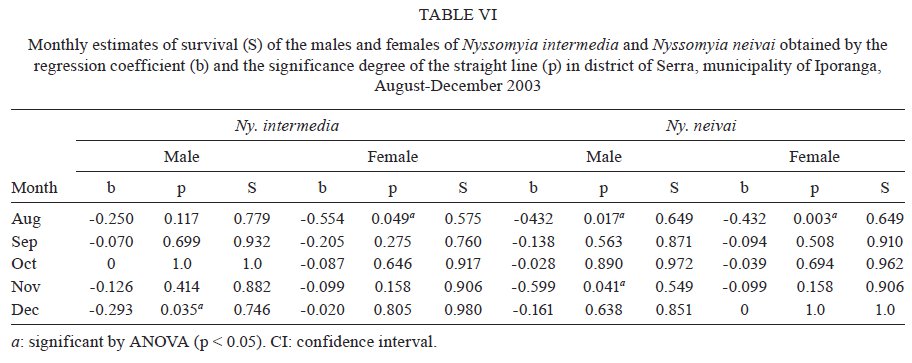
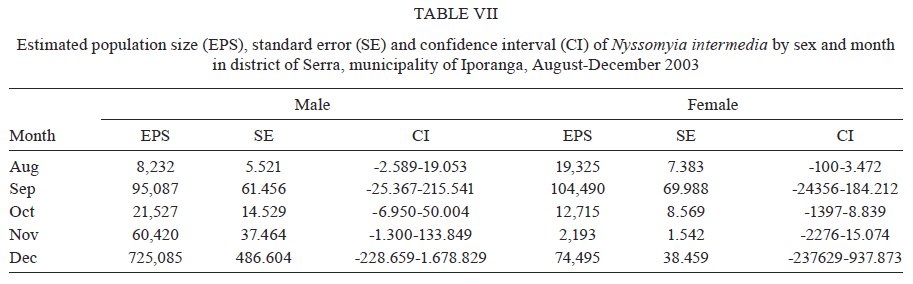
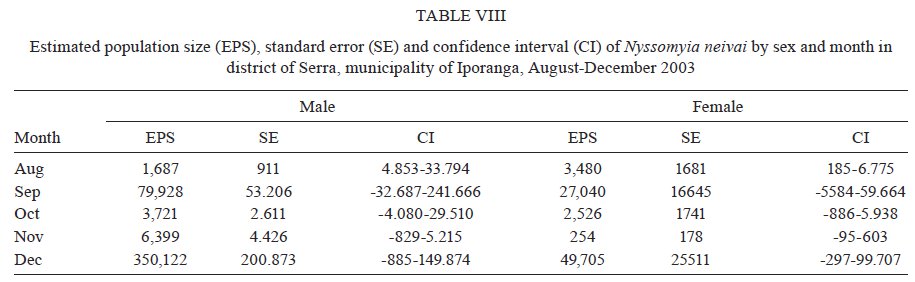
São Paulo, Av. Dr. Arnaldo 715, 01246-904 São Paulo, SP, Brasil Financial support: FAPESP (00/06811-0), IBAMA-SP (02027007000/2000-76) Received 19 February 2009 Code Number: oc09230 ABSTRACT The dispersal and survival of the phlebotomines Nyssomyia intermedia and Nyssomyia neivai (both implicated as vectors of the cutaneous leishmaniasis agent) in an endemic area was investigated using a capture-mark-release technique in five experiments from August-December 2003 in municipality of Iporanga, state of São Paulo, Brazil. A total of 1,749 males and 1,262 females of Ny. intermedia and 915 males and 411 females of Ny. neivai were marked and released during the five experiments. Recapture attempts were made using automatic light traps, aspiration in natural resting places and domestic animal shelters and Shannon traps. A total of 153 specimens (3.48%) were recaptured: 2.59% (78/3,011) for Ny. intermedia and 5.35% (71/1,326) for Ny. neivai. Both species were recaptured up to 144 h post-release, with the larger part of them recaptured within 48 h. The median dispersion distances for Ny. intermedia and Ny. neivai, respectively, were 109 m and 100 m. The greatest dispersal range of Ny. intermedia was 180 m, while for Ny. neivai one female was recaptured in a pasture at 250 m and another in a pigsty at 520 m, showing a tendency to disperse to more open areas. The daily survival rates calculated based on regressions of the numbers of marked insects recaptured on the six successive days after release were 0.746 for males and 0.575 for females of Ny. intermedia and 0.649 for both sexes of Ny. neivai. The size of the populations in the five months ranged from 8,332-725,085 for Ny. intermedia males, 2,193-104,490 for Ny. intermedia females, 1,687-350,122 for Ny. neivai males and 254-49,705 for Ny. neivai females. Key words: mark-release-recapture - sand fly - vectors - American cutaneous leishmaniasis Besides the vector competence of the insects, several other parameters, including those related to their relationship to the hosts of the agents and their survival, are considered in order to characterise their vectorial capacity (Reisen 1989). The dispersal of the insects is one of these parameters, as a clustered or random pattern of transmission of these agents depends in great part on the insects' mobility. For example, the studies of the dispersal of American sand flies have identified a maximum displacement of 960 m; the great majority, however, are recovered within 200 m (Chaniotis et al. 1974, Alexander 1987, Alexander & Young 1992) and 70 m (Casanova et al. 2005). About 10% of the almost 500 phlebotomine species described in the Americas have been implicated as vectors of Leishmania (CIPA Group 1999). The great majorities of these vectors are associated with forested areas and bite the humans who enter these environments or encounter humans when they move out towards neighbouring dwellings. The establishment of domestic animal populations in the peridomicile of these dwellings is common and may favour increase in the sand fly populations (Andrade Filho et al. 2001, Teodoro et al. 2003, Leonardo & Rebêlo 2004, Massafera et al. 2005). Nyssomyia intermedia (Lutz & Neiva, 1912) and Nyssomyia neivai (Pinto, 1926), with highly similar morphological and bio-ecological aspects, are suspected of being cutaneous leishmaniasis vectors in state of São Paulo (SP), Brazil (Andrade Filho et al. 2007). In this state, there are areas where these species are allopatric - Ny. intermedia being restricted to coastal areas and Ny. neivai to the interior plateau areas - while sympatric areas exist in the range that is transitional between coast and plateau (Marcondes et al. 1998, Andrade Filho et al. 2007). The Ribeira Valley region, with one of the highest cutaneous leishmaniasis incidence rates in SP, has a great part of its territory located in the sympatric range. The objective of this study was to undertake research into the dispersal and survival of these two probable vectors of cutaneous leishmaniasis in a rural area of the Ribeira Valley in the Serra District, municipality of Iporanga. MATERIALS AND METHODS Area studied - The research was undertaken in the Serra district (240 m asl) of the municipality of Iporanga (24º33'S, 48º40W) in the south of SP, Brazil. This district is situated in the speleological province of the Ribeira Valley, close to the Parque Estadual Turístico do Alto Ribeira (PETAR), an Atlantic forest reserve on the Serra de Paranapiacaba. It is located in a transi-tional area between the so-called Atlantic Plateau and the coastal plain. Its climate is also transitional between a tropical low altitude one and a temperate one of intermediate altitude (Nimer 1977) and is classified as sub-tropical and highly humid with no dry season. A coastal, hygrophilous, non-deciduous forest, with the highest trees attaining 25-30 m and many lianas, epiphytes, tree-ferns and palms (Karmann & Ferrari 2002) cover the non-anthropised areas. This district is situated on the banks of the Betari, a left bank tributary of the Ribeira do Iguape River. There are about 110 dwellings in the district (Giatti et al. 2004) distributed within a narrow strip along the road that links the cities of Iporanga and Apiaí and its economy is based mainly on catering to the tourists who visit the caves of the Santana nucleus, about 4 km away from the principal entrance of the Santana cave. Though it is an area of environmental preservation, it does not belong to the PETAR. Capturing, marking and releasing - Five experiments were undertaken, one per month, from August-December 2003. Each experiment lasted seven days. On the first day of each experiment, from 6-11 pm, sand flies were aspirated from Shannon traps or natural resting places under illumination by manual lanterns and then counted and placed in a box for marking with fluorescent dust. At the end of the collection period (11 pm), all the insects collected on that day were marked and released together at the same point as on all the other collection days in an area covered by bushy vegetation on the AG small farm (20 ha). For the recaptures, collec-tions were undertaken on the six successive days. The substrates and the Shannon traps where the insects were aspirated were all distributed within a range of 120 m from the release point. The box for marking the insects was constructed in accordance with the model of Casanova et al. (2005) with modifications: a sleeve was attached to the opening for the entry of the insects and the powder was blown into the box from the small lateral chamber. The fluorescent powder BioQuip® was used, with a different colour (yellow, green or red) utilized each month. The estimated numbers of insects of each species and sex that were marked and released were obtained as a proportion of the total of dead specimens that remained in the tag box after the releases. The identification of these dead insects was made in accordance with Galati (2003) after clarification using the method described by Forattini (1973). Recaptures - These were made on the AG small farm and on three other properties: the JRA smallholding (520 m south), the DO smallholding (180 m east) and in the Alambari de Baixo cave and the neighbouring forest (about 2 km northeast from the release site) in accordance with the scheme presented in Fig. 1. Fourteen fixed points were sampled with automatic light traps nightly from 18-6 h, except for those made in the ecotopes on the JRA smallholding, which were made on alternate nights. Diurnal (8-11 h) and nocturnal (19-22 h) aspirations were also undertaken daily at several sites on the AG small farm: on a rocky slope (diurnal, for about 1 h), in a rat hole (diurnal, sporadically), on tree trunks (nocturnal and diurnal, about 2 h per period), in pigsties (diurnal and nocturnal, about 1/2 h per period), in a hen-house and chicken nests (diurnal and nocturnal about 1/2 h per period), in a goose nest (nocturnal, sporadically) and, in September, in a burrow used by a bitch for its litter, situated among roots on the bank of a stream (nocturnal, 1/2 h). A Shannon trap was installed at the release point, but without regular periodicity during the six days post-release. A scheme of the ecotopes with their positions and distances in relation to the release point, as well as the technique of capture used, is presented in Fig. 1. The specimens were captured, separated by ecotope, technique and date and were then killed with chloroform and conditioned in Petri dishes, which were kept under refrigeration until the examination of the insects. This examination was made under a stereomicroscope coupled to a dark chamber and illuminated with ultraviolet light for fluorescence for the separation of tagged specimens. All the sand flies collected during recapture were submitted to clarification and identified as already mentioned. A sample of these specimens is deposited in the collection of the Departamento de Epidemiologia of the Faculdade de Saúde Pública, Universidade de São Paulo. The abbreviation of the name of each species follows the recommendations in Marcondes (2007). Statistical analysis - The χ2 or binomial test (the latter when each expected frequency was less than 5) was used to assess the different frequencies of the two species between distinct techniques, periods, sexes and ecotopes. Correlation analysis was undertaken between the proportion of insects recaptured and those released and between the median time and distances of recapture of the insects. The probability of association or correlation was considered at the level of p < 0.05. A semi parametric Cox regression survival analysis and a corresponding parametric one using an exponential model (constant lambda hazard rate of recapture) were undertaken using the total number of insects released and the numbers recaptured on the following days, according to the four species x sex combinations. Stata version 10 was the package used for analysis. A regression analysis was undertaken to estimate the daily survivorship. For this, the daily survival of males and females was considered constant throughout the time post-release and was calculated based on the recaptured insect numbers regressed and transformed by ln (y + 1), as a function of the days post-release (Nelson et al. 1978, Casanova et al. 2009): ln mt = ln (M0r) - t lnS, where mt is the number of marked insects recaptured on each day, M0 is the number of insects marked and released, r is the recapture rate, S is the daily survival rate and t is the time since release. The survival rate was calculated by the expression S = eb where e is the base of the natural logarithm and b the regression coefficient (Nelson et al. 1978) using the SPSS program for Windows. The sizes of the populations were calculated using the Lincoln index adjusted for low recapture rates with the inclusion of the survival in the estimate. The monthly population and variance were calculated by the average of the daily estimates in accordance with the formula Nt = (nt + 1) Mt / (mt + 1), being Mt = (M0 - " mt) St and the variance Var Nt = (Mt)2 (nt + 1) (nt - mt) / (mt + 1)2 (mt + 2) where Nt is the absolute population estimated, nt is the number of specimens captured, mt is the number of specimens marked and recaptured, Mt is the number of specimens marked that remained in the population, M0 is the number of specimens marked and released, S is the daily rate of survival and t is the time (days). The confidence intervals were calculated by the formula CI = average population ± 1.96 average standard error. RESULTS During the five experiments, a total of 6,310 sand flies were tagged, 28.3% of which died before they could be released, so a total of 4,396 specimens were released. Of this total, an estimated 3,011 (68.5%) were Ny. intermedia (1,749 males and 1,262 females), 1,326 (30.16%) were Ny. neivai (915 males and 411 females) and 1.3% were of four other species together: Pintomyia fischeri (1.07%), Migonemyia migonei (0.09%), Psathyromyia pascalei (0.16%) and Expapillata firmatoi (0.02%). Of the 4,396 specimens released, 153 (3.48%) were recaptured, 151 belonging to the genus Nyssomyia (98.7%) and two (1 of each gender) belonging to P. fischeri (1.3%), with both of these recaptured at the point of release. During the seven-day post-release captures, from August-December, a total of 73,259 specimens belonging to 20 sand fly species were captured. 69,916 (95.4%) of them belonged to Nyssomyia, of which 42,875 (61.3%) were Ny. intermedia and 27,036 (38.7%) were Ny. neivai (Table I). The numbers and percentages of insects recaptured were sorted by technique of capture and the proportions of Ny. intermedia and Ny. neivai collected by these techniques. The data are presented in Table I. The greatest numbers of marked insects of both species and sexes were obtained with automatic light traps. No males of either species were captured by Shannon trap nor were any females of Ny. neivai captured by nocturnal aspiration. As regards the proportion of marked specimens captured by each technique, the diurnal aspiration presented the highest returns for males of both species and for females of Ny. neivai, whereas the Shannon traps yielded the highest returns for the females of Ny. intermedia. The lowest returns with regards to both species and sex occurred with the automatic light traps. The numbers of specimens of Ny. intermedia and Ny. neivai sorted by site of recapture are presented in Table II. Tagged specimens of Ny. intermedia and/or Ny. neivai were recaptured at 17 sites, the former at 12 and the latter at 14. The maximum dispersal of Ny. intermedia was 180 m. Considering the two ecotopes of maximum range together, the females presented a greater and statistically more significant frequency than the males. For Ny. neivai, the maximum range was observed with one female recaptured at 520 m (Table II). The recapture occurred three nights after the release. The median dispersal distance of Ny. intermedia was 109 m (males 108 m and females 110 m) and that of Ny. neivai (males and females) 100 m. The proportion of recapture at distances of 100 m or more from the release point was 87.1% for males and 70.2% for females of Ny. intermedia and 71.4% for males and 62.1% for females of Ny. neivai. For Ny. intermedia, the highest recapture frequency (21.8%) was observed in a pigsty (100 m) and on the rock face (180 m) to the northwest; for Ny. neivai, the highest recapture frequency occurred in the same pigsty (22.5%) and in the hen-house/pigsty (18.3%). The high relative frequency, mainly of Ny. neivai, in the dog litter burrow is noteworthy, as the capture at this site was possible in only one month (September) due to the birth of the litter in that month. No tagged specimen was recaptured in the automatic light traps installed in forest environments, despite 353 specimens of Ny. intermedia and 154 of Ny. neivai having been captured in those traps installed at the edge of the forest (100 m from the release point) and 295 specimens of Ny. intermedia and 99 of Ny. neivai having been captured within the forest (100 m from the release point), whether in the forest or at the cave mouth (2 km from the release point), where 11 and 25 specimens of Ny. intermedia and 11 and 13 specimens of Ny. neivai were, respectively, captured. Fig. 2 presents the number of males and females of Ny. intermedia and Ny. neivai released and the percentage that were recaptured by month. The frequencies of the sexes released varied in the five experiments. Throughout the five experiments, a negative correlation was observed between the percentages of insects recaptured and the numbers released for both sexes of the two species, but this was statistically significant only for males of Ny. neivai (Ny. intermedia: males -0.56, females -0.40; Ny. neivai males -0.98, females -0.52). Both species were recaptured up to 144 h post-release, with the greater part of the specimens recaptured within 48 h. Ny. neivai presented the greatest and a statistically significant frequency for recapture prior to 24 h as compared with that of Ny. intermedia and the inverse result occurred at 120 h (Fig. 3). Males and females of Ny. intermedia had different peaks in their intervals of frequency, but the peaks were higher and statistically significant for females at 48 and 144 h and for males at 72 h. Both sexes of Ny. neivai presented their greatest frequencies prior to 24 h, without any significant differences between them, as occurred in the other intervals. The median values of the times at which the specimens of Ny. intermedia and Ny. neivai were recaptured by distance are presented in Table III. For Ny. intermedia, a negative correlation was observed between these two variables, but it was significant only when both sexes were taken into account. For Ny. neivai, the correlation was positive and significant for females and both sexes together. The recapture incidence rates for both sexes of Ny. intermedia and Ny. neivai, the numbers of insects released by sex, insect-time at risk during the observation period, insects recaptured and the mean hazard rate of recapture are presented in Table IV. The hazard ratios of recapture (by Cox survival analysis) with their confidence intervals for each sex of both species are shown in Table V. Though there are differences among these hazard rates, they are not statistically significant at the level of 95%. The estimated daily survival rate by month for males and females of the two species is presented in Table VI. For both sexes of Ny. intermedia and females of Ny. neivai, p was < 0.05 only in one of the months and for males of the latter species, p < 0.05 in two months, being the most significant in August. For the estimates of the size of the populations, the value of survival rate adopted for males and females of Ny. intermedia was, respectively, that of December (0.746) and August (0.575) and for both sexes of Ny. neivai the value of survival rate was that of August (0.649). The sizes of the estimated populations by sex of Ny. intermedia and Ny. neivai are presented, respectively, in Tables VII and VIII and the ratios of the estimated numbers of specimens of Ny. intermedia to Ny. neivai are shown in Table IX. In this latter table, the ratio of the insects released is also presented. The size of the populations presented a wide range of variation, including their confidence intervals. The most numerous populations of Ny. intermedia males and both sexes of Ny. neivai occurred in December and of Ny. intermedia females in September. Males and females of Ny. intermedia were, respectively, more numerous than those of Ny. neivai throughout the period, with the Ny. intermedia/Ny. neivai ratio varying between 1.19-9.44 for the males and 1.59-8.63 for the females. For the specimens released, these ratios follow the same tendency as those of the estimated post-release populations, but with a smaller range: males varied between 1.59-5.03 and females between 1.28-5.67. Though there was a clear predominance of Ny. intermedia over Ny. neivai, there is no significant difference between the populations of the two species by virtue of the great variances. DISCUSSION Dispersal values obtained in circumscripted samples of adult insects might be associated with their displacement for colonisation (fed and unfed females), their search for animal hosts (pre-fed females) or substrates for interbreeding (both sexes), their quest for resting places (both sexes) or their search for breeding places (fed females). Each one of these activities may be influenced, among other factors, by the fidelity that the insects present with regards to some substrates, as observed by Freitas et al. (2009) in relation to Ny. neivai, as well as by the capacity of the insects to respond to light stimuli in such a way as might result in distinct probabilities of capture depending on the techniques used. Furthermore, each insect's behavioural pattern might present distinct epidemiological significance. The dispersal pattern of Ny. neivai in the Mogi-Guaçu Valley, another endemic area for cutaneous leishmaniasis in SP has been investigated by Casanova et al. (2005), who reported an overall recapture rate of 7.5%. In regards to the specimens recaptured, the maximum respective range observed was 99 m for males and 128 m for females, with the arithmetical average distance being 53 m for the former and 55 m for the latter; 86% of the males and 89% of the females were recaptured at less than 70 m and 16% of them were recaptured in peridomiciliary habitats. The 84.3% of the specimens recaptured in Shannon traps situated at the edge of or within the forest led those authors to conclude focal dispersion. There was no previous information on Ny. intermedia. The recapture rates of Ny. intermedia (2.6%) and Ny. neivai (5.4%) observed in the present study were lower than that for this latter species obtained by Casanova et al. (2005) and Freitas et al. (2009), however, the maximum range for both sexes of Ny. intermedia (180 m) and for Ny. neivai (males 180 m and females 520 m) and the median dispersal distance for males (108 m) and for females (110 m) of Ny. intermedia and for males and females (100 m) of Ny. neivai were greater than those for Ny. neivai observed by those authors for Ny. neivai. The median values for both species were close, but below the average. Our data are closer to those obtained by Morrison et al. (1993) for Lutzomyia longipalpis in an endemic focus of visceral leishmaniasis in Colombia, where the recapture rates were 7.7% (males) and 1.5% (females), with 51% recaptured at 100 m or more. The greatest proportion of recapture in automatic light traps in our study also differs from that obtained by Casanova et al. (2005). This is perhaps due to the fact that we utilised fewer Shannon traps because this was the technique that produced the highest proportional result for females of Ny. intermedia and the second most productive result for Ny. neivai. Also different from those authors, only 4.2% of Ny. neivai specimens were captured on the rock face situated at the forest edge, though no single specimen was recaptured in the forest itself. However, Ny. intermedia had 21.8% of its tagged specimens aspirated from the rock face at the edge of the forest. These different frequencies for Ny. intermedia and Ny. neivai at the forest-edge may indicate a greater dependence of the former on the forest. On the other hand, the recapture of one female of Ny. neivai in a pasture at 270 m and another at 520 m in a pigsty in the peridomicile of the JRA smallholding (6 and 3 post-release days, respectively) seems to indicate the species' better adaptation to open and less humid environments, as in the interior area of the SP, where Ny. intermedia is absent and Ny. neivai occurs at very high frequencies (Casanova et al. 2005, Odorizzi & Galati 2007). Animals and humans seem to exercise great attraction for males, since 67.6% were recaptured in or near dwellings that were situated close to the release point; it is possible that the carbon dioxide and water vapour exhaled by the hosts and their odours, images and heat may stimulate the host-seeking response of sand flies, as identified for mosquitoes by McIver (1982) and presumably also for other nematocerous biting flies by Gibson & Torr (1999). On the other hand, this aggregation may contribute to high levels of pheromones being released by males, further attracting other specimens of this sex. The recapture of females in greater frequencies and at greater distances than the males (as observed at the sites representing the highest ranges of dispersal for Ny. intermedia, 180 m, where the females were statistically significantly more numerous than the males, with only females of Ny. neivai being captured at 270 m and 520 m) may be due to the search for sites for colonisation or blood sources. Consequently, they may help to disseminate the leishmaniasis agent that they might harbour from the enzootic foci in other areas, thus accounting for its domiciliary transmission in the Ribeira Valley (Gomes et al. 1986). The recapture risk of both sexes of Ny. neivai was higher than that of Ny. intermedia (according to the Cox survival model). This also means that the former species was recaptured faster than the latter, which can be explained by the lower survival rate of Ny. neivai. In fact, this lower survival rate was observed in the comparison specifically with the males of Ny. intermedia. So, in this case, the faster and higher recapture rates of Ny. neivai may at least partially explain why the activities related to the preservation of the species such as feeding, mating, looking for resting and breeding sites, tend to occur at a greater intensity and for shorter intervals for Ny. neivai than for Ny. intermedia and therefore increase the chances of the former's being caught. If this is true, a greater survival rate than that actually found for Ny. intermedia would also be expected. Although Andrade Filho et al. (2004) observed longer survival for Ny. intermedia than for Ny. neivai under laboratory conditions, in the present study the former presented a lower survival rate. Thus, perhaps its longer gonotrophic cycle (3-9 days with a peak on the 5th day) than that of Ny. neivai (4 days), as observed, respectively, by Rangel et al. (1985) and Casanova et al. (2009) means that the females remain in their resting places longer and their displacement in the search for hosts occurs at greater intervals, thus, their recapture risk is smaller. The majority of Ny. neivai specimens were recaptured within 24 h, which is different from the behaviour of those in the state of Paraná, observed by Freitas et al. (2009), and from those of Ny. intermedia in the present study, which were captured in greater numbers between 24-48 h. Ny. intermedia, beyond being recaptured more slowly than Ny. neivai, presented a negative correlation between the median time of recapture and the distance of the ecotopes. This suggests a faster displacement than that of Ny. neivai, for which this correlation was positive. This behaviour seems to give Ny. intermedia a greater potential for spreading pathogens. Casanova et al. (2009) inferred daily survival rates of 0.681 for males and 0.667 for females and a gonotrophic cycle of 4 days for an allopatric population of Ny. neivai in the Mogi-Guaçu Valley, also in SP. They also observed, under experimental conditions, that high infection levels by Leishmania braziliensis in the stomodeal valve of Ny. neivai occurred eight days post-infectious blood meals and at least 72 h after digestion and oviposition. Therefore, from the infectious blood meal, an interval of between 9-11 nights would be necessary for the transmission of the parasites, when only 1.2-2.6% of the potentially infectious females would still be alive. Thus, it may be concluded that the pattern of transmission of the cutaneous leishmaniasis with sporadic cases may be attributed to these circumstances. For Ny. intermedia, Rangel et al. (1992) observed massive experimental infection by L. braziliensis of the stomodeal valve 5-6 days after infectious blood meals. If this shorter extrinsic incubation period for L. braziliensis is also valid for the Ny. intermedia population investigated here, the greater size of the population of this latter species throughout the period of the experiment leads us to suppose that Ny. intermedia also has considerable participation in the transmission of the leishmaniasis parasite in the area even though its survival rate (0.565) was smaller than that of Ny. neivai (0.649). The low recapture rates obtained seem to be influenced by the great variance in the estimates of the population sizes in such a way as not to show any significant differences between them. Evidently, other parameters need to be assessed to obtain a more realistic picture of the role of the two species as vectors of the L. braziliensis in the area, mainly as regards their relationships with the hosts of the parasite and with the human population. Although a greater number of marked insects were recaptured with automatic light traps than by other techniques, their numbers as proportions of the total of those captured were very low (0.135% for males and 0.109% for females), meaning that the identification of the specimens gave a lot of work. The use of more Shannon traps would probably have increased the number of specimens, especially of females, recaptured, as occurred in the experiments of Casanova et al. (2005, 2009), though carried out under greater constraints. The results of nocturnal aspirations in the recapture attempts show male/female ratios of 10.3:1.0 and 8.9:1.0, respectively, for Ny. intermedia and Ny. neivai; during the morning aspirations, these ratios were lower: 3.3:1.0 and 2.9:1.0. These differences in the diurnal and nocturnal aspirations may be due to many females that took blood during the night but stayed in the ecotope, mainly in the pigsty during the morning (it was possible for us to observe this), while others were being aspirated from natural resting places. At night, while the females go in search of a blood source, the males wait to copulate with them, hence the high frequencies of this latter sex on tree trunks or other substrates such as fence posts, perhaps attracted by released pheromones. The alternation in the frequencies of males and females in the captures of insects before being tagged and released during the five experiments (Fig. 2) seems to be an intrinsic feature of these species, as monthly collections from 2001-2003 in the same place observed on several occasions and is probably not significantly correlated with rainfall or temperature (Galati et al. 2009, unpublished observations). The negative correlation observed between the proportion of insects recaptured and those released may be influenced by the small numbers of the latter. Thus, as the denominator is small, the fewer insects recaptured, the greater the possible distortion of the resulting values of the proportion. Another hypothesis arises from the fact that when the insects are very numerous in the box for marking, they become more stressed and tends to stay longer in their resting places after being released, which could not be taken into consideration in the sample proportions. The small number of specimens recaptured provides only limited information on the potential distance that the insects may traverse, as the maximum distance registered for the recapture of marked insects depends on the limits of the sampling (Alexander & Maroli 2003). However, the marking, releasing and recapturing studies help to determine the spatial range of dispersal, the understanding of which may make it possible to reduce the contact between humans and insects and, consequently, control the spread of the leishmaniasis agent. Therefore, although our studies present the same limitations as those cited by other authors and the displacement of Ny. neivai at distances much greater than those observed by Casanova et al. (2005) has been demonstrated, the great majority of the marked insects were found to be restricted to a range of 180 m from the release point. It is possible that the presence of many domestic animals acts as a factor limiting dispersal and that, in areas without them, other patterns may be found. However, despite the presence of the domestic animals, the dispersal of both species to the veranda of a neighbouring dwelling as well as to another peridomicile where the tree in which the trap was hung was very close to the house, shows a certain tendency to frequent human habitats, probably due in part to the electric lighting in these environments and also to human kairomones at the sites of recapture. As in the locality studied, the dwellings are situated between the Betari River and the surrounding slopes covered with dense vegetation and within the known dispersal range of both species, so the occurrence of epidemic outbreaks of leishmaniasis is explicable. Furthermore, the dispersal pattern found for the two sand fly species and their high frequencies in the area studied also help us to understand the high average annual coefficients of incidence. The coefficient of incidence of 152.9/100,000 inhabitants for this disease in the municipality of Iporanga between 1998-2006 (CVE 2007) is one of the highest in SP, probably because this environmental feature is frequent throughout the region. ACKNOWLEDGEMENTS To the guide, Jânio Ribeiro de Andrade, and his family, to Mr. Antônio Gurgel, owners of the two small farms, for permitting us to make the captures on their properties, to Maria Dulce Bianchi Rosa and Adriana Yurika Maeda, for helping us in the selection of the sand flies, to Paloma Helena Shimabokuru, for the organization of the data base, to Aristides Fernandes, for helping us with some of the experiments, Dr. Ademir Galati, for providing the necessary logistic support, to Prof. José Maria Pacheco de Sousa, for helping us in the survival analysis, to the anonymous reviewers, for their suggestions, and to Mr. Arthur Anthony Boorne, for his correction of the English text. REFERENCES
Copyright © 2009 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc09230t5.jpg] [oc09230t1.jpg] [oc09230f3.jpg] [oc09230t6.jpg] [oc09230t2.jpg] [oc09230t7.jpg] [oc09230f1.jpg] [oc09230f2.jpg] [oc09230t8.jpg] [oc09230t3.jpg] [oc09230t4.jpg] [oc09230t9.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}