 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 104, No. 8, 2009, pp. 1165-1170 ARTICLES Ecological niche modelling and differentiation between Rhodnius neglectus Lent, 1954 and Rhodnius nasutus Stål, 1859 (Hemiptera: Reduviidae: Triatominae) in Brazil Taíza Almeida BatistaI; Rodrigo Gurgel-GonçalvesII, + ILaboratório
de Zoologia, Curso de Ciências Biológicas, Universidade Católica
de Brasília, Brasília, DF, Brasil Financial support: FAP-DF, CNPq Received 10 August 2009 Code Number: oc09232 ABSTRACT Ecological niche
modelling was used to predict the potential geographical distribution of Rhodnius
nasutus Stål and Rhodnius neglectus Lent, in Brazil and to
investigate the niche divergence between these morphologically similar triatomine
species. The distribution of R. neglectus covered mainly the cerrado
of Central Brazil, but the prediction maps also revealed its occurrence in transitional
areas within the caatinga, Pantanal and Amazon biomes. The potential distribution
of R. nasutus covered the Northeastern Region of Brazil in the semi-arid
caatinga and the Maranhão babaçu forests. Clear ecological
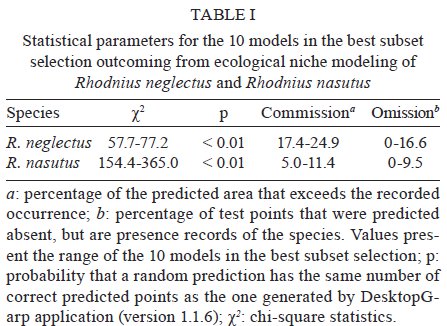
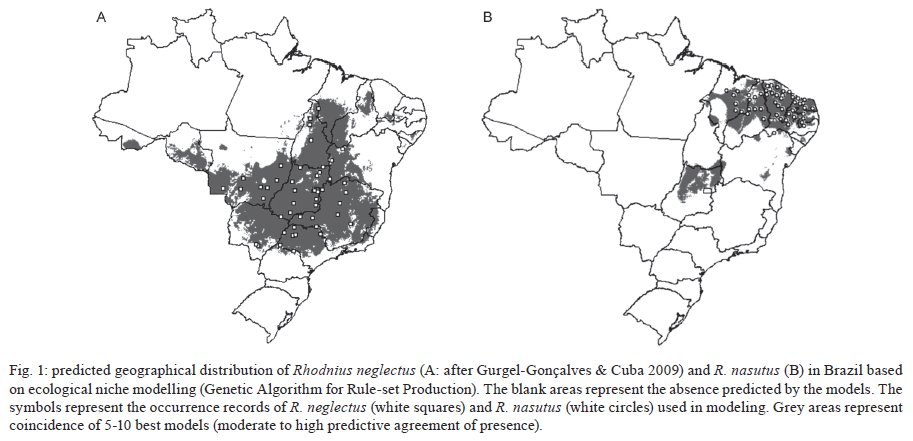
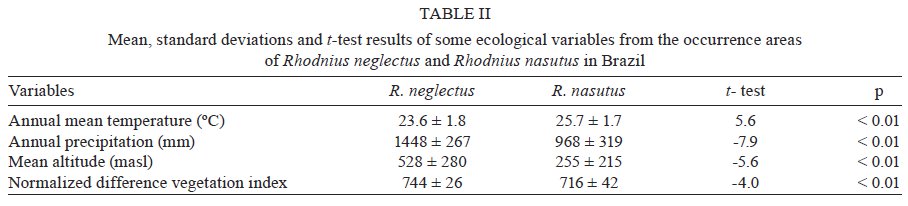
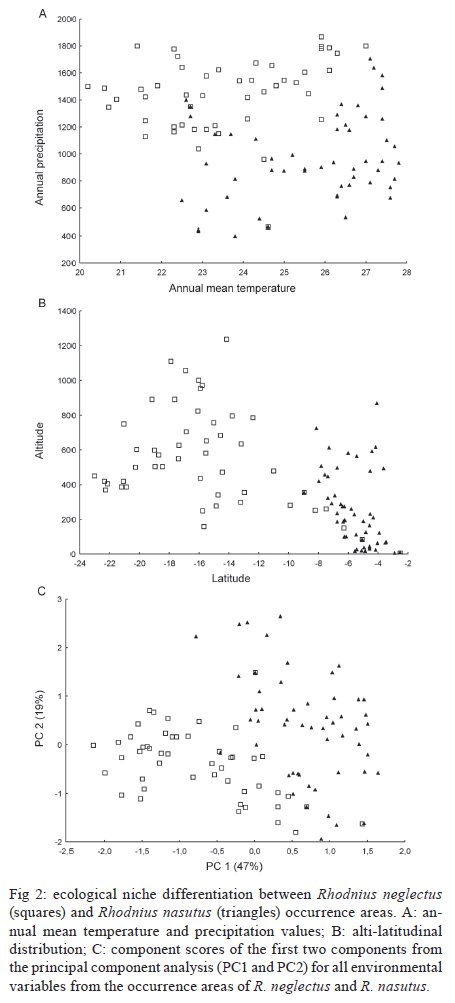
niche differences between these species were observed. Key words: Rhodnius neglectus - Rhodnius nasutus - GARP - geographical distribution - ecological niche modelling - Brazil Ecological niche modelling (ENM) is a methodological approach that generates geographical distribution models for species. It has been a useful tool in studies on zoogeographical distribution patterns (Anderson et al. 2003, Tsoar et al. 2007). In vector studies, ENM suggests potential areas of vector occurrence and consequently, risk areas of pathogen transmission (Peterson 2006). This approach has already been used to analyse the geographical distribution of some triatomine species (Costa et al. 2002, Peterson et al. 2002, López-Cárdenas et al. 2005, Gurgel-Gonçalves & Cuba 2009), thus contributing to epidemiological studies on Chagas disease. Rhodnius neglectus Lent, 1954 is a common (although non-endemic) species of the cerrado biome in Central Brazil (Gurgel-Gonçalves & Cuba 2009). Rhodnius nasutus Stål, 1859 seems to be restricted to the Northeastern Region of Brazil in the semi-arid caatinga, including the states of Ceará (CE), Maranhão (MA), Pernambuco (PE), Piauí and Rio Grande do Norte (Lucena & Lima-Borba 1977, Carcavallo et al. 1999, Galvão et al. 2003). These triatomines are frequently found in palm trees and bird nests in sylvatic environments, but adult specimens infected by Trypanosoma cruzi Chagas, 1909 have been invading houses in Central and Northeastern Brazil (Sarquis et al. 2004, Gurgel-Gonçalves et al. 2008). The problem of reinfestation of insecticide-treated dwellings highlights the need for a better understanding of the geographical distribution and ecology of wild vector species (Guhl et al. 2009). Thus, predicting the distribution of these triatomines may improve vector surveillance strategies for Chagas disease. R. neglectus and R. nasutus are morphologically similar triatomines, distributed in open formations with long dry seasons in Central and Northeastern Brazil, respectively. Although R. neglectus occurs mainly in the cerrado and R. nasutus in the drier caatinga, there is evidence that these two species may occur sympatrically in transitional areas between their respective core ecoregions (Abad-Franch et al. 2009). However, the geographical distribution limits of these species are still poorly known. Thus, the aims of our study were: (i) to predict the potential geographical distribution of R. nasutus and R. neglectus in Brazil using the ENM approach, (ii) to investigate the ecological niche differences between these species, (iii) to identify the environmental variables that most influence their geographical distribution and (iv) to identify potential areas of co-occurrence for R. nasutus and R. neglectus. MATERIALS AND METHODS Occurrence records for R. nasutus and R. neglectus - The distributional data for R. nasutus were obtained from Lent (1948), Lent (1954), Lucena and Lima-Borba (1977), Lent and Wygodzinsky (1979), Silveira et al. (1984), Pinto and Bento (1986), Carcavallo et al. (1999), Galvão et al. (2003), Sarquis et al. (2004), Dias et al. (2008), Lima and Sarquis (2008) and Abad-Franch et al. (2009). Distributional data for R. neglectus were obtained from Gurgel-Gonçalves and Cuba (2009). Geographical coordinates of the municipality administrative centre were obtained from the Brazilian Institute of Geography and Statistics - IBGE (http://www.ibge.gov.br) database. All maps were created and edited using the software ARC VIEW (version 3.2). ENM and niche differentiation - Among the ENM methods developed, the Genetic Algorithm for Rule-set Production (GARP) (Stockwell & Peters 1999) has demonstrated its utility for predicting species distributions (Anderson et al. 2003). Twenty-one variables were used to generate models using the DesktopGarp application (version 1.1.6). Details about the environmental database and the software parameters used in GARP are described elsewhere (Gurgel-Gonçalves & Cuba 2009). The χ2 statistic was used to calculate the probability of a random prediction being similar as the one generated by GARP. For each species we produced 100 models. The best subset procedure (Anderson et al. 2003) was used to select 10 of 100 models with the highest predictive values. These models were imported into ARC VIEW (version 3.2) and summed using the Map Calculator function in order to generate a single cumulative predictive map ranging from zero (predicted absence) to 10 (the coincidence of the 10 best models, the highest predictive agreement of presence). We considered the coincidence of the 5-10 best models (moderate to high predictive agreement of presence) to be appropriate for describing the species' potential distribution. This procedure added a component of conservatism to the predictions of GARP, which otherwise could over-extrapolate the potential areas. We used Student's t-test to compare the mean altitude, temperature, normalised difference vegetation index (NDVI) and precipitation in the areas of occurrence of R. neglectus and R. nasutus (alpha = 0.01). Furthermore, we conducted a principal component analysis (PCA) with all 21 environmental variables from the occurrence areas of R. neglectus and R. nasutus. PCA reduces the dimensionality of the original set of variables with little loss of information by transforming the original variables into a new set of independent components (Foottit & Sorensen 1992). The components that accounted for the majority of the total variance were examined and the most highly loaded variables were analysed. This approach was used to identify which environmental variables most influence the geographical distribution of these species. A scatterplot of the component scores of the species occurrence areas was created to visualise the ecological niche differences between R. neglectus and R. nasutus. PCA and Student's t-test were computed with StatisticaÒ (StatSoft Inc, Tulsa, OK, USA). RESULTS We analysed 49 occurrence records for R. neglectus and 54 for R. nasutus. The 10 best GARP models resulting from ecological niche analysis for these species presented statistically significant χ2 values (p < 0.01), indicating that the models are quite predictive and that they summarise the ecological requirements of each species. The highest levels of commission and omission errors were observed for R. neglectus (Table I). The predicted distribution of R. neglectus covered mainly the cerrado of Central Brazil, but the models also revealed moderate to high predictive agreement of presence of this species in transitional areas, such as cerrado-caatinga [western Bahia (BA)], cerrado-Pantanal [Mato Grosso do Sul (MS)], cerrado-Amazon (northern areas of Mato Grosso and Tocantins), cerrado-Paraná interior forests [western São Paulo (SP)] and cerrado-Maranhão babaçu forests (north of MA and PI), as shown in Fig. 1A. The potential distribution of R. nasutus covered the Northeastern Region of Brazil in the semi-arid caatinga and the Maranhão babaçu forests. However, the prediction maps of R. nasutus indicated areas of potential occurrence for this species in Central Brazil (Fig. 1B). ENM of R. neglectus and R. nasutus supports potential areas of co-occurrence for these species in the Maranhão babaçu forests and in caatinga/cerrado transitional areas, mainly in PI (Fig. 1A, B). However, these species showed statistically different values for the environmental variables of their occurrence areas (Table II). Clear ecological niche differences between these species were observed. R. nasutus occurred more in warmer and drier areas than R. neglectus (Fig. 2A). Moreover, the occurrence areas of R. neglectus showed the highest values of altitude and NDVI (Table II, Fig. 2B). In the multivariate analysis, including all environmental variables from the occurrence areas for R. neglectus and R. nasutus, the first two principal components (PC) summarised 66% of the environmental variance and also showed clear niche differences between these species (Fig. 2C). PC1 explained most of the total variation and was correlated with altitude and temperature variables, mainly temperature in the coldest and driest months. PC2 explained 19% of the environmental variance and was correlated with NDVI and precipitation variables (Table III). Low values of precipitation/NDVI combined with high values of temperature in the coldest and driest months distinguish the ecological niche of R. nasutus from that of R. neglectus. DISCUSSION The use of the ENM approach to analyse the predicted geographical distribution of triatomines has been increasing in recent years (Costa et al. 2002, Gorla 2002, Peterson et al. 2002, Rodriguero & Gorla 2004, López-Cárdenas et al. 2005, Sandoval-Ruiz et al. 2008, Arboleda et al. 2009, Gurgel-Gonçalves & Cuba 2009, Gurgel-Gonçalves & Silva 2009, Ibarra-Cerdeña et al. 2009). These predictions have yielded ecological and biogeographical knowledge regarding triatomines that may lead to new vector-control and surveillance strategies for Chagas disease. The geographical distribution of R. nasutus reviewed in the present study included occurrence records from PE and BA and was wider than that recorded by Carcavallo et al. (1999). We also updated the alti-latitudinal distribution and described the ecological niche for this species. The association between R. nasutus and palm tree species may also explain the distribution patterns observed. R. nasutus occurs mainly in the carnauba wax palm Copernicia prunifera (Mill.) HE Moore, where it feeds mainly on birds (Lent & Wygodzinsky 1979, Pinto & Bento 1986, Carcavallo et al. 1998, Sarquis et al. 2004). However, R. nasutus inhabits other palm tree species such as Attalea speciosa Mart. and Syagrus oleracea (Mart.) Becc., which have a wide distribution in the caatinga and cerrado biomes (Bento et al. 1992, Dias et al. 2008). Moreover, the colonisation of this triatomine in another tree species (Licania rigida Benth.) was reported by Lima and Sarquis (2008). These authors also suggest that environmental damage is facilitating the ability of R. nasutus to colonise other trees besides palms. The potential distributions of these tree species should be investigated and compared with R. nasutus' distribution in future studies to better understand these associations. The predicted distribution of R. neglectus was similar to the one presented by Gurgel-Gonçalves and Cuba (2009), but in the present study we increased the range of the predicted area in Southwestern Brazil after including records from MS and SP. Although R. neglectus occurs mainly in the cerrado biome and R. nasutus in the drier caatinga, potential distribution areas of R. neglectus in the caatinga and R. nasutus in the cerrado were observed. The presence of R. neglectus in caatinga is already registered (Abad-Franch et al. 2009, Gurgel-Gonçalves & Cuba 2009), but available distributional data do not show the presence of R. nasutus in the core area of the cerrado; all known records of Rhodnius in this region are due to R. neglectus (Lent & Wygodzinsky 1979, Carcavallo et al. 1999, Galvão et al. 2003, Gurgel-Gonçalves et al. 2003, 2004, 2008). According to Anderson et al. (2003), this overprediction error in ENM approaches derives from potentially habitable regions correctly predicted as presence, but that cannot be demonstrated as such because there is no occurrence record of the species there. Indeed, the predicted areas of R. nasutus in Central Brazil (mainly in state of Goiás) should be related to low precipitation values in the dry season. The absence of R. nasutus in Central Brazil may have various causes, including historical restrictions (e.g., geographic barriers and/or lack of sufficient dispersal opportunities) and biotic interactions (such as competition with related species; e.g., R. neglectus). By examining the congruence or discord-ance between predicted and actual distributions, it is possible to evaluate the potential role of ecological and historical factors in determining a species' geographical distribution (Anderson et al. 2002). The temperature, altitude, NDVI and precipitation values of R. neglectus and R. nasutus occurrence areas were compared and our results showed clear ecological niche differences between these species. PCA was also useful for measuring niche amplitude for each species, as previously reported for North American triatomines (Ibarra-Cerdeña et al. 2009). The maximum variation in niche amplitude occurred along the first axis (PC1), mainly associated with a temperature gradient. Moreover, a precipitation-humidity gradient was observed along the second axis (PC2). Our results support the hypothesis that the warmest and driest areas of Northeastern Brazil produce ideal conditions for R. nasutus occurrence. In contrast, the coldest areas with the highest altitude and NDVI values seem to be favourable for R. neglectus. Other studies addressing the influence of climatic variables on triatomine distribution have also showed ecological niche differences between triatomine species (Costa et al. 2002, Sandoval-Ruiz et al. 2008, Arboleda et al. 2009). For example, the vapour pressure deficit minimum was the principal variable among the nine most important to describe the distribution of Rhodnius pallescens, which is in agreement with its stenohydric status (Arboleda et al. 2009). Thus, the distribution of Rhodnius species may be explained by various climatic factors. Apparently, the level of humidity is one of the main factors separating species of humid environments (most species of Rhodnius) from those of dry environments, such as R. neglectus and especially R. nasutus. Further studies analysing the influence of environmental variables on the geographical distribution of all 16 known Rhodnius species should reinforce this hypothesis. Finally, these studies should have implications for the surveillance of Chagas disease vectors, because there is evidence that triatomine household infestation is restricted to the driest regions (Abad-Franch & Monteiro 2007, Sandoval Ruiz et al. 2008). ENM of closely related species occurring in adjoining or slightly overlapping areas is useful for exploring the factors affecting the species' geographical distributions and for providing directional hypotheses that can be tested in future studies. Investigations at smaller spatial scales on the ecology of these species (e.g., microhabitat, feeding patterns), especially comparisons among populations occurring inside and outside the predicted overlap zone, should give conclusive results on the roles of competitive interactions and historical factors in shaping species distributions (Costa et al. 2008). Further comparative phylogeographical studies may also evaluate the role of demography and climatic events in shaping the diversity and distribution of R. neglectus and R. nasutus populations in South America. In Brazil, the predicted maps support potential areas of co-occurrence for these species in MA, PI, CE, PE, BA, state of Paraíba (PB) and state of Alagoas. There are occurrence records of R. neglectus and R. nasutus sharing ecotopes in the same localities in PI (Bento et al. 1992) and BA (Abad-Franch et al. 2009). The same pattern may occur in other areas including the MA, PI, CE, PE and PB that have favourable macroenvironmental conditions for both species to occur. Such areas should be investigated to broadly compare the realised distribution with the one predicted by the models and to identify microenvironmental factors segregating these species. Entomologists engaged in Chagas disease vector surveillance should be aware that the morphologically similar triatomines R. neglectus and R. nasutus can occur in the same localities of Northeastern Brazil, as predicted in our study. Thus, the identification of bugs in these areas should be improved by applying morphometrical (Gurgel-Gonçalves et al. 2008) and/or molecular (Abad-Franch & Monteiro 2005) methods. ACKNOWLEDGEMENTS To Ricardo B Machado, for providing the database compiled for analysis using GARP, to Karen Schmidt, for assistance in earlier methodological analysis, to Fábio Oliveira Alves, for great fieldwork assistance, and to César A Cuba Cuba, Fernando Abad-Franch, Fernando A Monteiro, Liléia Diotaiuti, Lourdes MA El-moor, Loureiro and Renato Caparroz, for suggestions on an preliminary versions of this manuscript. REFERENCES
Copyright © 2009 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc09232t1.jpg] [oc09232f1.jpg] [oc09232t3.jpg] [oc09232t2.jpg] [oc09232f2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}