 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 104, No. 8, 2009, pp. 1171-1176 ARTICLES Bromeliad-inhabiting mosquitoes in an urban botanical garden of dengue endemic Rio de Janeiro - Are bromeliads productive habitats for the invasive vectors Aedes aegypti and Aedes albopictus? Márcio Goulart MocellinI; Taynãna César SimõesII; Teresa Fernandes Silva do NascimentoI; Maria Lucia França TeixeiraIII; Leon Philip LounibosIV; Ricardo Lourenço de OliveiraI, + ILaboratório
de Transmissores de Hematozoários, Instituto Oswaldo Cruz-Fiocruz, Av.
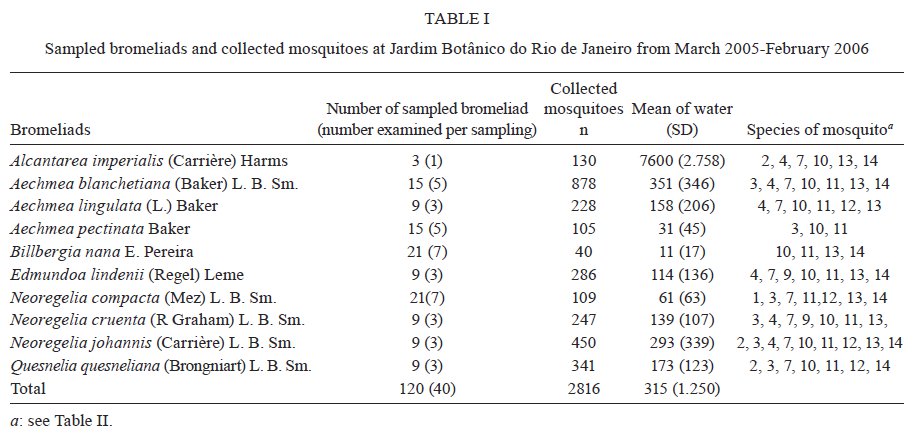
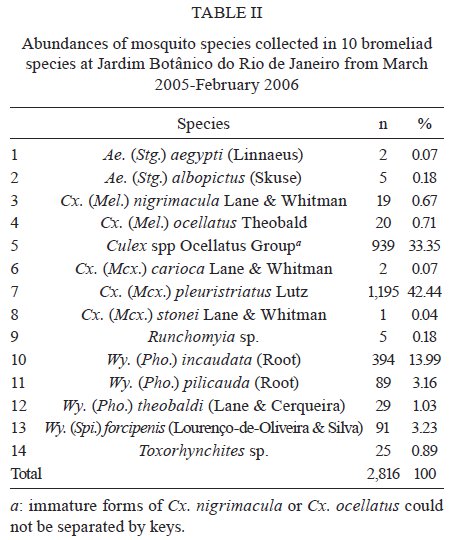
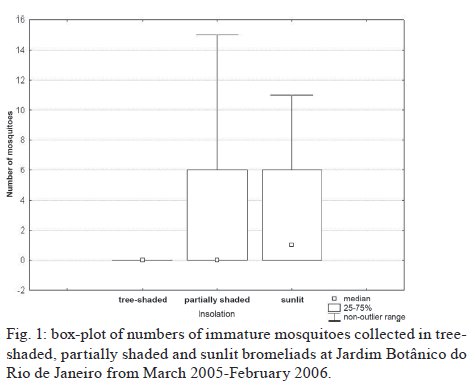
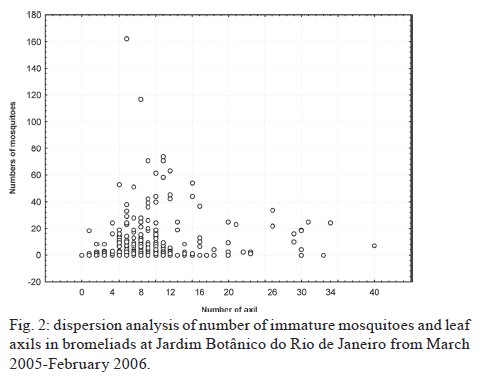
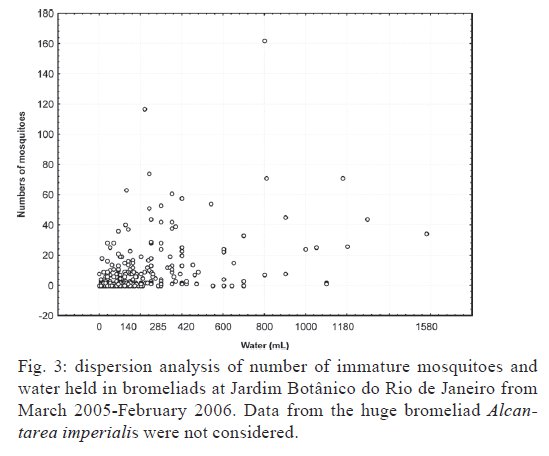
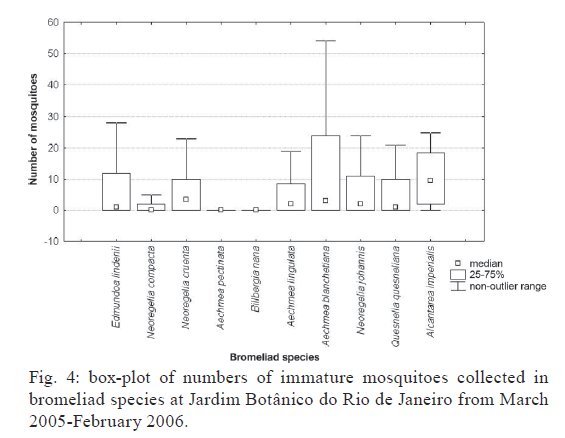
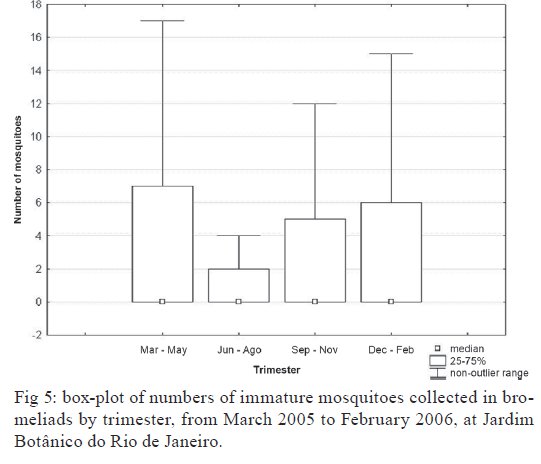
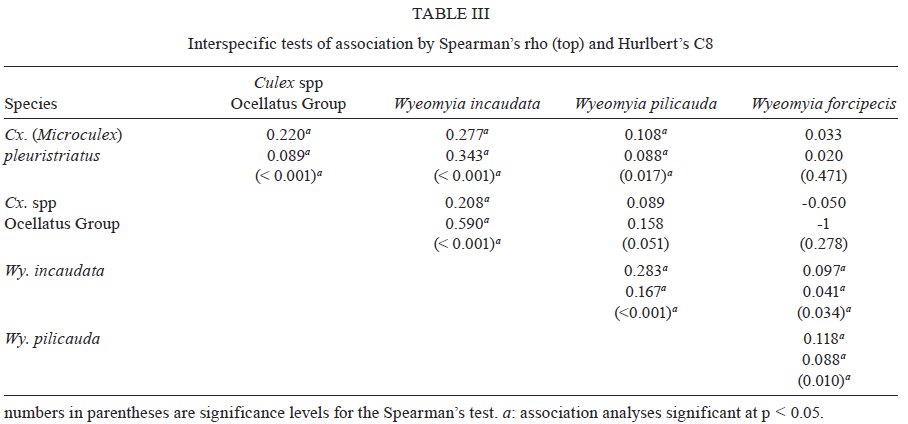
Brasil 4365, 21045-900 Rio de Janeiro, RJ, Brasil Financial support: CNPq (501529/2003-9) Received 30 October 2009 Code Number: oc09233 ABSTRACT Immatures of both Aedes aegypti and Aedes albopictus have been found in water-holding bromeliad axils in Brazil. Removal of these plants or their treatment with insecticides in public and private gardens have been undertaken during dengue outbreaks in Brazil despite uncertainty as to their importance as productive habitats for dengue vectors. From March 2005-February 2006, we sampled 120 randomly selected bromeliads belonging to 10 species in a public garden less than 200 m from houses in a dengue-endemic neighborhood in Rio de Janeiro. A total of 2,816 mosquito larvae and pupae was collected, with an average of 5.87 immatures per plant per collection. Culex (Microculex) pleuristriatus and Culex spp of the Ocellatus Group were the most abundant culicid species, found in all species of bromeliads; next in relative abundance were species of the genus Wyeomyia. Only two individuals of Ae. aegypti (0.07%) and five of Ae. albopictus(0.18%) were collected from bromeliads. By contrast, immatures of Ae. aegypti were found in manmade containers in nearly 5% of nearby houses. These results demonstrate that bromeliads are not important producers of Ae. aegypti and Ae. albopictus and, hence, should not be a focus for dengue control. However, the results of this study of only one year in a single area may not represent outcomes in other urban localities where bromeliads, Ae. aegypti and dengue coincide in more disturbed habitats. Key words: Aedes aegypti - Aedes albopictus - dengue - bromeliad mosquitoes - Bromeliaceae Although domestic Aedes aegypti has a clear preference for man-made larval habitats, its immatures may be found in natural containers such as water-containing axils of bromeliads growing in both modified and semi-modified environments of the Americas (Peryassú 1908, Forattini & Marques 2000, Marques et al. 2001, Cunha et al. 2002, O'Meara et al. 2003, Fouque et al. 2004, Varejão et al. 2005, Maciel-de-Freitas et al. 2007). Bromeliads are popular ornamental plants and often used in public and private gardens. The abundance of bromeliads where dengue is endemic has been construed to represent a threat to dengue vector control (Natal et al. 1997, Forattini et al. 1998, Forattini & Marques 2000). Even though Ae. aegypti immatures have been found in such plants only occasionally (Cunha et al. 2002, O'Meara et al. 2003, Maciel-de-Freitas et al. 2007, David et al. 2009) and few studies have evaluated the productivity of ornamental bromeliads for Ae. aegypti (Frank et al. 1988) compared to other larval habitats, bromeliads have been arbitrarily insecticide-treated, removed or incinerated during some dengue epidemics, such as in outbreaks in Rio de Janeiro (RJ) in 2001-2002 and 2008 (Lourenço-de-Oliveira 2008, JR Duarte, unpublished observations). Herein we describe results of field surveys of the identity and frequency of bromeliad-inhabiting mosquitoes in bromeliads in a public garden in a dengue-endemic area of RJ and demonstrate that native species of Culex and Wyeomyia mosquitoes were the most abundant culicids and that the invasive species Ae. aegypti and Aedes albopictus were recovered rarely from these plants, which should not be of concern for dengue control. MATERIALS AND METHODS One hundred and twenty bromeliads belonging to 10 species cultivated in the ground at a public garden [Jardim Botânico do Rio de Janeiro (JBRJ); 22°58'04"S 43°13'44"W] were randomly selected, labeled and numbered. The selected bromeliads were growing in the open, less than 200 m from houses in the dengue-endemic neighborhood of Gávea. One of the borders of JBRJ is an ecotone between the garden environment and secondary rain forest, but the sampled bromeliads were located at the other edge, in a modified environment where the garden is contiguous to houses. The bromeliads were exposed to natural rainfall and were not treated with insecticide during the sampling period nor in the previous 10 months. Labeled bromeliads were divided into three groups, each composed of an equal number of specimens belonging to a single species. The number of specimens per group (Table I) varied according to the population density of the bromeliad species at JBRJ. From March 2005-February 2006, bromeliads of all groups were sampled every three months. For instance, bromeliads from group one were sampled in March, June, September and December 2005, while sampling from those of group two began in April 2005, and so on. At each sampling, all the water impounded by all the leaf axils of a labeled bromeliad was aspirated according to Lozovei and Silva (1999), measured (mL) and taken to the laboratory for mosquito identifications using keys for morphological characters of immature stages or emerged adults provided by Lane and Whitman (1951), Corrêa and Ramalho (1956), Coutinho and Forattini (1962), Forattini and Toda (1966), Cotrim and Galati (1977), Consoli and Lourenço-de-Oliveira (1994) and Motta and Lourenço-de-Oliveira (2005). The number of axils holding water and whether the sampled plant was in the sun or partial or total shade was recorded at every sampling. Daily rainfall was obtained from the meteorological station at JBRJ. For statistical analyses we used the programs SPSS, Minitab and Statistica 6.0. For purposes of correlation analyses, samples from the same plant separated by three months were considered as independent. Spearman's correlations (significance level: p < 0.05) were used to evaluate relationships between total mosquito abundance and (i) the number of bromeliad leaf axils holding water and (ii) the total amount of water (mL). The same test was applied to calculate the correlations between abundances of individual mosquito species and (i) the amount of water held in the bromeliad's leaf axils and (ii) rainfall. Nonparametric Kruskal-Wallis tests (p < 0.05) were used to analyze variations in mosquito abundance in each bromeliad species, in relation to their location in open-sunlit, shaded or partially shaded sites and amount of water held in each sunlit bromeliad. Ae. aegypti and Ae. albopictus were not considered in statistical analyses because of their rarity in collections. A Friedman's test was used to analyze effects of bromeliad location (sun light vs. shade) in which bromeliad species was considered a block effect. RESULTS AND DISCUSSION A total of 2,816 immature mosquitoes was collected. Species of Culex (77.2%) and Wyeomyia (21.4%) were the most abundant bromeliad mosquitoes (Table II). Culex (Microculex) pleuristriatus was by far the most common species (42.5%), followed by species of Culex belonging to the Ocellatus Group (34.7%). Only two larvae of Ae. aegypti (0.07% of total mosquitoes) and five of Ae. albopictus (0.18%) were collected in more than one year of sampling of the bromeliads. This result suggests that larvae of these species are inferior competitors with native bromeliad-inhabiting species or that adult females prefer not to oviposit in bromeliads. Manmade containers (e.g., drains, plant vases, plastic basins, tires, water tanks) in nearby houses were consistently positive for both species of Aedes, particularly Ae. aegypti. The house infestation index for Ae. aegypti nearby was found to be as high as 5% in a survey conducted simultaneous to our sampling of bromeliads in 2006 (Secretaria Municipal de Saúde, unpublished observations). During the 2001 and 2008 dengue epidemics in RJ, it was hypothesized that bromeliads in public gardens and even native bromeliads growing in the ecotone between the natural and modified environments would be productive and persistent Ae. aegypti habitats from which emerged adults would invade houses and recolonize manmade larval habitats. Our findings suggest just that this concern is unfounded, as only two larvae and no pupae of Ae. aegypti were found in a single bromeliad compared to high infestations in surrounding houses. Varejão et al. (2005) did not detect any relationship between Ae. aegypti occurrence in native bromeliads on rock slopes and the Stegomyia indices in adjacent areas in the state of Espírito Santo, Brazil. Marques et al. (2001) found more Ae. albopictus larvae in exotic urban bromeliads than in native or periurban ones in the state of São Paulo and considered bromeliads to be of secondary importance as larval habitats for Ae. albopictus compared to artificial containers. The number of collected immatures and species composition varied according to bromeliad species (Table I). Around 31% of all mosquitoes were found in Aechmea blanchetiana, Neoregelia compacta, Neoregelia cruenta and Neoregelia johannis, which showed the greatest diversity of mosquito species composition in their axils. Species of Wyeomyia were the most frequent mosquitoes in N. compacta (68.8%) and Billbergia nana (50%). With mosquito species designated as blocks, a Friedman's test indicated a marginally significant effect of bromeliad location on mosquito abundance (χ2 = 5.64, DF = 2, p = 0.06) (Fig. 1). However, Lopez (1997) (apud Lopez et al. 1998) have suggested that shade favors faunal abundance, species composition and richness in the leaf axils of bromeliads. The number of immature mosquitoes collected per sampled bromeliad was positively correlated with the number of axils per bromeliad (Spearman's ρ = 0.234, p < 0.0001, n = 480) (Fig. 2) as well as with the amount of water held by the plant (ρ = 0.529, p < 0.0001, n = 468) (Fig. 3). When considering mosquito species separately, positive correlations were found between the amount of water in the plant and the number immatures of: Wyeomyia incaudata (ρ = 0.495, p < 0.0001, n = 480), Cx. pleuristriatus (ρ = 0.438, p < 0.0001, n = 480), Wyeomyia pilicauda (ρ = 0.280, p < 0.0001, n = 480), Culex ocellatus (ρ = 0.238, p < 0.0001, n = 480), Culex nigrimacula (ρ = 0.193, p < 0.0001, n = 480), Wyeomyia theobaldi (ρ = 0.149, p = 0.001, n = 480) and Wyeomyia forcipenis (ρ = 0.136, p = 0.003, n = 480). The number of mosquitoes found in the huge bromeliad Alcantarea imperialis was not considered for these analyses, because mosquito numbers differed from those in the other plants and the amount of water held by A. imperialis was usually 10-20 folds greater than that held by the other species (Table I). The median number of mosquitoes collected in A. imperialis was among the highest (Fig. 4) reported among bromeliad species and the number of mosquitoes collected per sample varied much less in this huge bromeliad species than in most of other plants, such as A. blanchetiana and species of genus Neoregelia (Fig. 4). Although rainfall reported in the first two trimesters (Mar-Aug) of the sampling period was lower than in the third and fourth trimesters (Sept-Feb), the number of mosquitoes per collection in all bromeliads did not differ significantly among trimesters (Kruskal-Wallis H = 3.17, p = 0.366) (Fig. 5). Correlations between lagged rainfall and mosquito abundances were analyzed, following Lourenço-de-Oliveira et al. (2004). Significant correlations (n = 480, p < 0.05) were observed between rainfall in the first week before collections for abundances of Cx. nigrimacula (ρ = 0.630) and Wy. pilicauda (ρ = 0.585). The five most common mosquito species occurred together in the same sample significantly more often than predicted by chance (Table III). The absence of significant negative correlations in the associations of species suggests that females of these species are using similar clues for oviposition in favorable bromeliads. Species of the genus Wyeomyia and Culex have frequently been found together in bromeliads (Marques et al. 2001, Yanoviak et al. 2006). Interestingly, the frequency of these Culex species in the cultivated bromeliads at JBRJ is similar to values reported in native plants in the natural environment elsewhere in Brazil (Corrêa & Ramalho 1956, Lourenço-de-Oliveira et al. 1986, Marques et al. 2001, Silva et al. 2004). Some mosquitoes, such as Culex (Microculex) and Culex belonging to the Ocellatus Group and some Wyeomyia (Phoniomyia) species seem to be intimately associated with bromeliads in the Neotropical region. Bromeliad-specific Wyeomyia spp have been shown in Florida to inhibit the growth and survivorship of Ae. albopictus co-occurring in axils of Bilbergia pyramidalis (Lounibos et al. 2003). Interspecific competition with bromeliad-specialist species may make this habitat less suitable for the container generalist species Ae. albopictus and Ae. aegypti in JBRJ. Our results and previous works (Marques et al. 2001, Cunha et al. 2002, Maciel-de-Freitas et al. 2007) lead to the conclusion that these bromeliads are not important or even productive microhabitats for Ae. aegypti and, hence, do not need to be treated for dengue control. Additionally, we hypothesized that the occasional findings and frequency of Ae. aegypti in bromeliads in the modified environment may be correlated to high infestation levels in such an area as result of competition for ovipositing habitats. However the mosquito fauna in bromeliads maintained domestically in yards or houses, where Culex spp and Wyeomyia spp may be less common than in JBRJ, deserves to be investigated, because bromeliads have became popular houseplants in dengue endemic areas. Indeed, the results of this study of only one year in a single area may not represent outcomes in other urban localities where bromeliads, Ae. aegypti and dengue coincide in more disturbed habitats. Also, it is known that the suitability of tank bromeliads for Ae. albopictus depends on bromeliad shape, e.g., Neoregelia spp being more suitable for development than Billbergia spp (Raban 2006). Moreover, our results may not apply to areas where bromeliads are not endemic yet are grown as ornamental plants in private gardens, such as Hawaii, where Ae. albopictus flourishes in bromeliad axils in the absence of competing native mosquito species (Yang et al. 2003). AKNOWLEDGEMENTS To Jair Rosa Duarte, for the suggestions in the design of this investigation, to Bruno Rezende Silva, for the bromeliads identification, to Marcelo Quintela Gomes, Kleber Soares de Sousa, Mauro Menezes Muniz, Marcelo Celestino dos Santos, Reginaldo da Silva Rego e Renato Carvalho de Andrade, for the support in the field work. REFERENCES
Copyright © 2009 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc09233f3.jpg] [oc09233t1.jpg] [oc09233f5.jpg] [oc09233t3.jpg] [oc09233f1.jpg] [oc09233t2.jpg] [oc09233f2.jpg] [oc09233f4.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}