 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
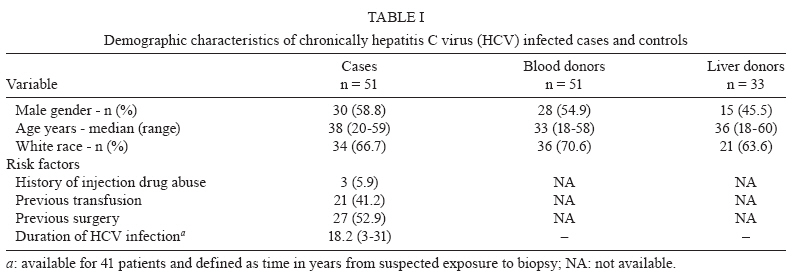
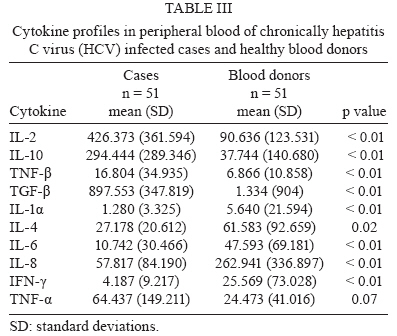
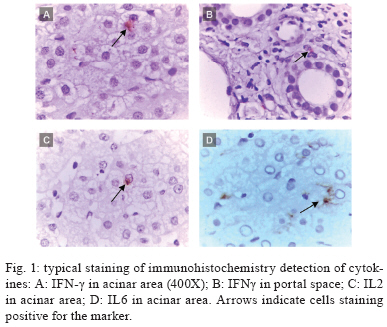
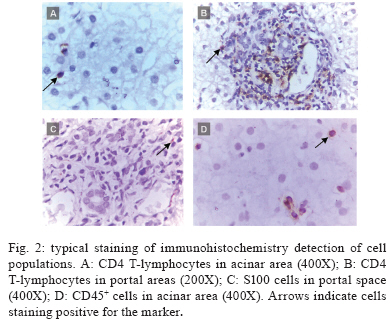
Memórias do Instituto Oswaldo Cruz, Vol. 105, No. 1, 2010, pp. 25-32 ARTICLES Tissue and serum immune response in chronic hepatitis C with mild histological lesions AT R-VisoI, +; MIS DuarteII; C PagliariII; ER FernandesII; RA BrasilII; G BenardIII; CC RomanoIII; S OgusukuIII; NP CavalheiroI; CE MeloI; AA BaroneI ILaboratório
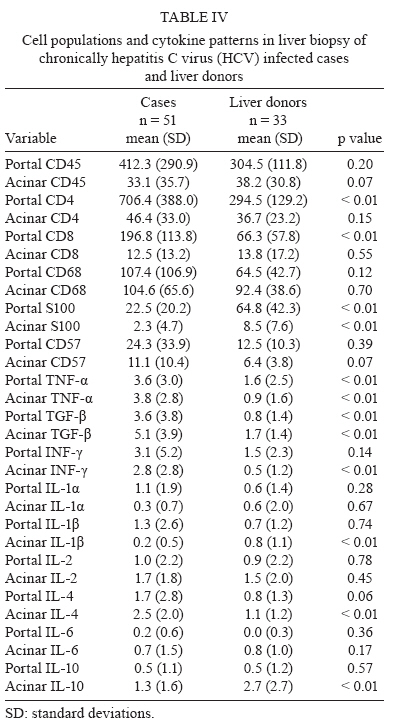
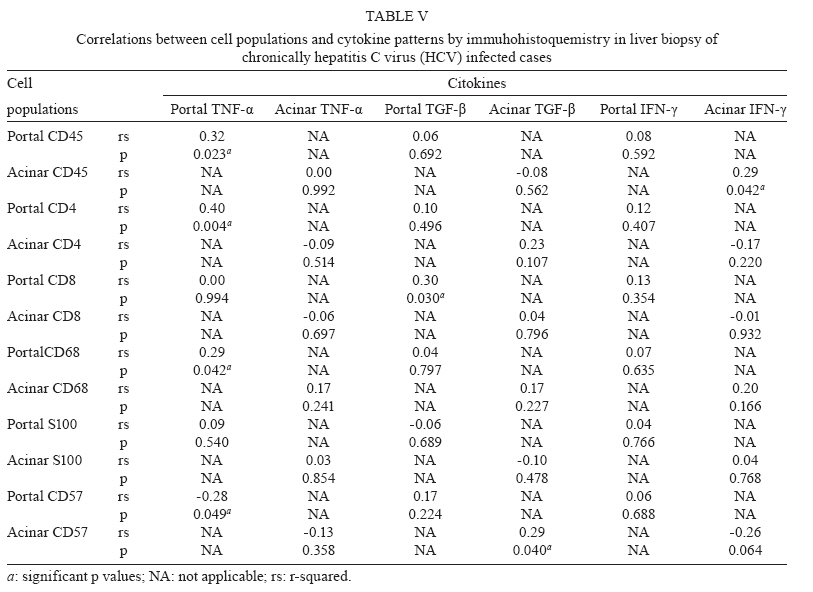
de Hepatite Financial support: FAPESP (00/14345-0) Received 28 July 2009 Code Number: oc10003 ABSTRACT The immunopathogenesis of chronic hepatitis C virus (HCV) infection is a matter of great controversy and has been suggested to involve a complex balance between cytokines with pro and anti-inflammatory activity. We investigated the expression of inflammatory cells and cytokines in the liver and serum of 51 chronically HCV infected patients and compared them to data from two sets of normal controls: 51 healthy blood donors and 33 liver biopsies of healthy liver donors. We also assessed the relationship between selected cytokines and cell populations in hepatic compartments and the disease stage. Compared with controls, hepatitis C patients had a greater expression of portal TNF-α, TGF-β and CD4+ and acinar IFN-γ, TNF-α, IL-1β and IL-4, as well as a higher serum concentration of IL-2, IL-10 and TGF-β. Significant positive correlations were found between portal CD4+ and TNF-α, portal CD8+ and TGF-β, portal CD45+RO and TNF-α, acinar CD45+RO and IFN-γ and acinar CD57+ and TGF-β. In conclusion, we have shown that (i) in this sample of predominantly mild disease, the immune response was associated with a pro-inflammatory response pattern, (ii) CD4+ T-lymphocytes played a major role in orchestrating the immune response and (iii) these events primarily took place in the portal space. Key words: chronic hepatitis C - cytokines - immunohistochemistry - physiopathology of chronic hepatitis C - T-lymphocytes Hepatitis C virus (HCV) is a major cause of chronic liver disease, with around 130 million people infected worldwide, accounting for an estimated 27% of cirrhosis cases and 25% of primary haepatocellular carcinoma cases (Alter 2007). Both viral and host immune mechanisms are involved in chronic infections, with the spontaneous resolution of HCV linked to vigorous and multi-specific T-cell responses and to attenuated CD4+ and CD8+ cell responses associated with viral persistence (Chang et al. 2001, Thimme et al. 2001, Day et al. 2002). Several studies have suggested that T-cell immunoregulatory cytokines play a key role in both HCV viral persistence and in the extent of liver damage. Therefore, while some cytokines may exert a pro-inflammatory activity, such as IL-1 (α and β), IFN-α, IL-8, TNF-α and IL-2, which can prime T-cells towards a Th-1 type immunity, others have a predominantly anti-inflammatory activity, as is the case for IL-4, IL-6 and IL-10, which are involved in Th-2 immunity. Additionally, some of these cytokines may have a fibrogenic (e.g., TGF-β) or an anti-fibrogenic (e.g., IFN-α) role (Tilg et al. 2006). Currently, it is unclear whether several inflammatory processes are the result of pure Th-1 or Th-2 type responses, or, as in hepatitis C, both arms (Th1 + Th2) are involved (Brown & Neuman 2001). Nevertheless, the expression of some cytokines is more closely related to the severity of the disease. IFN-γ is clearly augmented in the serum of chronic hepatitis C patients (Tilg et al. 1992, Cacciarelli et al. 1996, Kamal et al. 2004) and its increase has been correlated with an increase in the severity of disease (Gonzalez-Peralta et al. 1994). Moreover, an increased expression of IL-1, IL-2, IL-6, IL-10 and IL-18 has been documented in either the blood or liver of chronically HCV infected patients (Makris et al. 1994, Schvoerer et al. 2003, Hassan et al. 1997, Oyanagi et al. 1999, Polyak et al. 2001). Nonetheless, most studies have been limited by a reduced sample size, resulting in a lack of statistical power. Additionally, the great variability in the type of patients that were included in the studies, the techniques employed for cytokine assessment and the source of the specimens (peripheral blood or liver) has made it difficult to compare their results. In this study, we aimed to investigate the immune response to HCV through expression of inflammatory cells and cytokines in the liver and serum of 51 chronically HCV-infected patients compared to normal controls. We further assessed the relationship between the selected cytokines and cell populations in both the peripheral and hepatic compartments with the stage of disease. PATIENTS AND METHODS Patients and controls - Patients were consecutively recruited from the Infectious Diseases Division at the University Hospital from March 2001-October 2002. Most of the patients were referred from the blood bank service of the hospital. All of the patients underwent a complete medical and laboratory evaluation and a liver biopsy. The inclusion criteria for this study was as follows: (i) an age between 18-60 years, (ii) a confirmed HCV infection (by a second or third-generation ELISA or immunoblot technique), (iii) a positive qualitative PCR for HCV-RNA, (iv) a 1.54-4.00 fold increase in serum alanine aminotransferase (ALT) and (v) a liver biopsy confirming chronic hepatitis, graded and staged according to Ishak et al. (1995). Patients with any other hepatic diseases, previous immunosuppressive treatment, HIV or human T cell lymphotropic virus infection, or concurrent malignant disease were excluded. Two sets of controls were selected for this study; healthy blood donors were selected for cytokine pattern comparisons and liver biopsy specimens from the organ donors were used as controls for immunohistochemistry features. The institutional review board approved the study and all of the patients and blood donors provided written informed consent. Virological markers - Quantification of HCV-RNA was performed with an Amplicor HCV Monitor (Roche Diagnostic Systems, Mannheim, Germany) at a lower detection limit of 600 IU/mL. Genotyping was carried out by using the purified product of the nested-PCR, as delimited by NCR4 primers. The samples were submitted to sequencing (ABI Prism DNA Sequencer) using the ABI Prism Big Dye Terminator Cycle Sequencing Ready Reaction version 2.0 (Applied Biosystems). The sequences were then analysed and compared with compatible genotypes within the HCV Sequence Data Base with BLAST (Basic Local Alignment Search Tool, 2003). Tissue sample processing - The biopsy specimens, obtained during a routine diagnosis of the patients at the University Hospital, were fixed in 10% formalin and embedded in paraffin for light microscopy. Only the biopsy specimens measuring at least 1 cm or bearing at least 10 portal spaces were analyzed. The specimens were then submitted to haematoxylin-eosin staining, as well as Masson's trichromate, reticulin and Perls' techniques. Two experienced histopathologists independently performed all of the analyses. The case specimens were then graded and staged according to the semi-quantitative Ishak score (Ishak et al. 1995). Immunohistochemistry - Expression of cytokines in the biopsy specimens of HCV-cases and liver donor controls was semi-quantitatively assessed through immunomarking by immunohistochemistry, following the streptavidin-biotin-peroxidase immunohistochemical method of detection. The following specific monoclonal antibodies were employed: IFN-γ (MAB285), TNF-α (AF-210-NA), IL-1α (AF-200-NA), IL-1-β (AF-201-NA), IL-2 (AF-202-NA), IL-4 (AF-204-NA), IL-6 [AF-206-NA and IL-10 (MAB217) (all from R&D Systems)] and transforming growth factor (TGF)-β (MCA 797; SEROTEC). Cell subpopulations were assessed with the following specific monoclonal antibodies: mouse anti-CD45+RO (M742), mouse anti-CD4 (M834), mouse anti-CD8 (M7103), mouse anti-CD68 (M876), rabbit anti-S100 (Z311) (all from DAKO) and mouse anti-CD57 (1166 CH; IMMUNOTECH). The sections were dewaxed in xylene and hydrated through a graded ethanol series. After this, 3% hydrogen peroxide was used to block endogenous peroxidase. The sections were incubated overnight with primary mouse antibodies and the streptavidin-biotin-peroxidase immunohistochemical method of detection was used. The Catalysed Signal Amplification-CSA, based on the peroxidase catalysed deposition of a biotinylated phenolic compound, was used to better visualise the positive cells. Biotinylated anti-mouse antibody was applied to the sample for 15 min at 37ºC, following application of the primary antibodies. Peroxidase-labelled streptavidin-biotin complex was then applied for 15 min at 37ºC, followed by the biotinyl tyramide reagent for 15 min and a secondary reaction with streptavidin peroxidase. The slides were counterstained with Harris haematoxylin and 3.3 diamino-benzidine tetrahydrochloride (SIGMA Chemical Company) was used as chromogen. Counting of the cytokines and cell populations was performed with an optical microscope and a square optical grid (1 cm2). Screening began at the first portal space on the left upper extremity and then moved to the right and downwards, with the grid sequentially centred on each portal space and the surrounding area. Within each screening square, positive cells were counted in the portal and the periportal spaces at a 400X magnification. At least 5-10 portal spaces were counted on each slide. The number of positive cells was divided by the number of counted spaces and this result was divided by 0.0625 (Weibel 1979). Cytokine assay - A quantitative measurement of serum cytokines was performed for the cases and blood donor controls using a commercially available enzyme-linked immunosorbent assay (R&D Systems), according to the manufacturer's instructions. The following cytokines were assessed: IL-1α, IL-2, IL-4, IL-6, IL8, IL-10, TNF-α, TNF-β, IFN-γ and TGF-β. The minimum detection limit was 10 pg/mL; thus, any measures below this threshold were considered to be zero. Statistical analysis - For descriptive analyses, the means and standard deviations, or the medians and interquartile ranges, are reported. Comparisons of proportions between the cases and controls were assessed by Chi-square test or Fisher's exact test, when appropriate. Continuous and ordinal data were compared by Mann-Whitney rank sum test. The Spearman's rank correlation coefficient was used to assess correlations between the continuous variables. A significance level of 0.05 was used and all of the p values are two-sided. All of the analyses were conducted in SPSS version 12 (2002). RESULTS Characteristics of the cases and controls - Fifty-one HCV-patients were included in the study and compared with 51 healthy blood donors and 33 liver donors. The demographic and epidemiological characteristics of the cases and controls are shown in Table I. Thirty (58.8%) cases were male, with a median age of 38 years (range 20-59). The cases and controls did not differ significantly with regard to gender, age or race. For 41 cases, the mean disease duration from exposure to biopsy was available and was 18.2 years (range 3-31). The detailed laboratory and histological findings of the cases are listed in Table II. In summary, the median aspartate aminotransferase and ALT levels were 45 IU/L (range 30-197) and 69 IU/L (range 32-399), respectively; 11 patients (21.5%) had hypoalbuminaemia and 16 patients (31.4%) had hypergammaglobulinaemia. The majority of patients (76.5%) were infected with HCV genotype 1 and 59.2% had an HCV RNA viral load greater than 850.000 IU/mL. The median Ishak score for histology was 5 (range 3-14) and a minimal and mild disease accounted for 78.4% of the pathologic findings (31.4% and 47.1%, respectively). Confluent hepatocyte necrosis was not seen. Grade I fibrosis (F1) was seen in 30 cases (58.8%) and portal inflammation was found in 29 (56.8%) cases. All of the cases had Councilman bodies in the hepatic parenchyma and the hepatic sinusoids presented with mild hyperplasia and hypertrophy in Kupffer cells, with a predominance of lymphocyte infiltrates. The majority (78%) of cases showed some evidence of steatosis, which was either mild, moderate or severe, either focused or generalized and either macro or microvacuolar. Biopsies from the liver donor biopsies showed no evidence of pathologic findings. A mild haemosiderin deposition and glycogenic degeneration was seen in one and two samples, respectively. Cytokine patterns in the cases and blood donor controls - Compared with the blood donor controls (Table III), the cases had significantly higher serum levels of IL-2 (426 ± 362 vs. 91 ± 124 pg/mL; p < 0.01), IL-10 (294 ± 289 vs. 38 ± 141 pg/mL; p < 0.01), TNF-β (17 ± 35 vs. 7 ± 11 pg/mL; p < 0.01) and TGF-β (898 ± 348 vs. 1 ± 904 pg/mL; p < 0.01). On the other hand, the cases showed significant lower levels of the following cytokines: IL-4 (27 ± 21 vs. 62 ± 93 pg/mL; p = 0.02), IL-6 (11 ± 30 vs. 48 ± 69 pg/mL; p < 0.01), IL-8 (58 ± 84 vs. 263 ± 337 pg/mL; p < 0.01) and IFN-γ (4 ± 9 vs. 26 ± 73 pg/mL; p < 0.01). Cell populations and cytokine patterns in liver biopsies of the cases and liver donor controls - In immunohistochemistry reactions, the positive cytokines and cell populations were stained brown, as shown in Figs 1, 2, respectively. The cell populations from the cases had a higher expression of portal CD4+ (706.4 ± 388.0 vs. 294.5 ± 129.2 cells/mm2; p < 0.01) and CD8+ (196.8 ± 113.8 vs. 66.3 ± 57.8 cells/mm2; p < 0.01) than the controls (Table IV). The cytokine patterns of the cases had a higher expression of acinar IL-4 (2.5 ± 2.0 vs. 1.1 ± 1.2 cells/mm2; p < 0.01), TNF-α (3.8 ± 2.8 vs. 0.9 ± 1.6 cells/mm2; p < 0.01), IFN-γ (3.1 ± 5.2 vs. 1.5 ± 2.3 cells/mm2; p < 0.01) and TGF-β (5.1 ± 3.9 vs. 1.7 ± 1.4 cells/mm2; p < 0.01), compared to the controls. The cases also expressed significantly higher portal TNF-α (3.6 ± 3.0 vs. 1.6 ± 2.5 cells/mm2; p < 0.01) and TGF-β (3.6 ± 3.8 vs. 0.8 ± 1.4 cells/mm2; p < 0.01) than the controls. The cases showed a significantly lower expression of portal S100 dendritic cells (DC) (22.5 ± 20.2 vs. 64.8 ± 42.3 cells/mm2; p < 0.01), ac-inar S100 (2.3 ± 4.7 vs. 8.5 ± 7.6 cells/mm2; p < 0.01), ac-inar IL-1β (0.2 ± 0.5 vs. 0.8 ± 1.1 cells/mm2; p < 0.01) and acinar IL-10 (1.3 ± 1.6 vs. 2.7 ± 2.7 cells/mm2; p < 0.01). Correlation patterns of cell population and cytokines among cases - The correlations between the cell markers and cytokines was analysed at either the portal spaces or the acinar areas (Table V). Significant positive correlations were seen between the following measures at the portal spaces: CD45+RO and TNF-α, CD4+ and TNF-α, CD8+ and TGF-β, CD68+ and TNF-α, CD57+ and IL-10, S-100 and IL-10. At the acinar level, CD45+ and IFN-γ, CD45+ and IL-1α, CD57+ and TGF-β and CD57+ and IL-2 were positively correlated, while CD68+ and IL-6 and CD68+ and IL-10 were negatively correlated. The correlations between the degree of liver damage and the cell populations and cytokines were also assessed. In immunohistochemistry analyses of the cases, periportal activity was positively correlated with portal CD4+ expression (r = 0.4; p < 0.01), while focal inflammation was positively correlated with acinar CD4+ (0.33; p = 0.02). A significant positive correlation for cytokine density was found between periportal hepatitis and portal IL-1β (r = 0.36; p < 0.01) and between acinar activity and acinar TGF-β (r = 0.34; p = 0.02). Following a correlation analysis of the serum cytokines and the degree of liver lesions, IL-8 was positively correlated with fibrosis (r = 0.29; p = 0.04) and IL1-α was negatively correlated with focal inflammation (r = -0.29; p = 0.04); the other serum cytokines did not show any correlation with liver compromised liver (data not shown). DISCUSSION In this study, we systematically assessed the role of a comprehensive array of cytokines and cell populations in chronically HCV infected patients and compared them with normal controls. Among the cases, we found that there was a greater expression of portal TNF-α, TGF-β, CD4+ and CD8+ and acinar IFN-γ, TNF-α, TGF-β and IL-4, as well as a higher serum concentration of IL-2, IL-10 and TGF-β compared to the controls. Cytokines are key mediators of inflammation, apoptosis, necrosis and fibrosis and they are actively involved in the regeneration process of liver tissue after injury. It has been hypothesised that successful treatment of hepatitis C depends on a complex balance between pro and anti-inflammatory responses (Bertoletti et al. 1997, Tsai et al. 2003, Wan et al. 2009). In our study, we found both pro and anti-inflammatory responses, with a predominant pro-inflammatory response pattern in the liver mediated by IFN-γ and TNF-α and modulated by CD4+, CD8+ and CD45RO+ T lymphocytes. This complex balance is further supported by the significant correlations found between: (i) portal CD4+ and TNF-α, (ii) portal CD8+ and TGF-β, (ii) portal CD45+RO and TNF-α and (iv) acinar CD45+RO and IFN-γ. The latter correlations that involve CD45+RO suggest that there is an ongoing activation of the cellular immune response, which is in agreement with the study by Murata et al. (2002). In line with other reports (Minutello et al. 1993, Sakai et al. 1999, Exley et al. 2002), our results demonstrate the compartmentalised nature of the immune response to HCV. The correlation between periportal activity and portal CD4+, as seen in a preliminary study (Viso et al. 2007) and the increased expression of portal TNF-α and acinar IFN-γ, TNF-α and IL-4, which was not replicated in serum, suggests that there was an intra-hepatic response. Apparently, most of the immune response occurred at the portal space level, supported by the lack of correlation between the hepatic and serum cytokines among the cases. TNF-α production is an early event in the pathogenesis of liver damage, triggering the synthesis of other cytokines. It is required for the proliferation of normal hepatocytes in liver regeneration, but it also mediates hepatocyte death (Zhu et al. 1998). The increased expression of portal and acinar TNF-α and acinar IFN-γ in the cases compared to the controls reinforces the major role of these pro-inflammatory cytokines in the liver disease caused by HCV. IL-2 is considered to be a Th1 type cytokine and is involved in enhancing the proliferation and activation of most T-lymphocytes, NK cells and B-lymphocytes. Liver sinusoidal and inflammatory cells have been reported to be sources of IL-2 and no consensus exists on the predictive value of this cytokine. Apparently, expression of IL-2 is associated with a more advanced stage of disease, as previously reported (Makris et al. 1994, Napoli et al. 1996, Bozkaya et al. 2000, Gramenzi et al. 2005). In our sample, although a higher concentration of IL-2 was found in the serum, no significant expression was found in the liver from the cases compared to the normal controls. This could possibly be due to the relatively mild disease that was observed in most of our cases, agreeing with reports from other authors (Woitas et al. 1997, Falasca et al. 2006). Indeed, it has been hypothesised that failure to secrete IL-2 might lead to a disruption of IFN-γ and proliferative capacity, contributing to the development of a persistent infection (Semmo et al. 2005). Our findings agree with those of Gramenzi et al. (2005), who reported that there was an inverse correlation between ALT levels and IL-2 expression when cytokine profiles of patients with chronic disease were compared with patients who had persistently normal serum ALT levels. IL-6 is a major mediator of the inflammatory process in the acute phase response, acting as a blocker of cell apoptosis (Baumann & Gauldie 1990) and promoting the differentiation of naïve T-cells into Th1 cells. IL-6 has been associated with disease progression and has been reported to be elevated in both hepatic and peripheral compartments (Malaguarnera et al. 1997, Oyanagi et al. 1999, Lapinski et al. 2001, Falaska et al. 2006). Interestingly, in this study, IL-6 was not associated with either chronic HCV infection or with the stage of disease. Our results are in contrast to those of other researches, but agree with those of Cribier et al. (1998), who found similar patterns in serum IL-6 between HCV-infected and non-infected patients. Although, it should be noted that IL-6 was above the cut-off value in only 11 patients, possibly due to the overall lower severity of disease sample. However, these findings deserve further investigation with appropriately powered studies. TGF-β has been implicated as a mediator of hepatic fibrogenesis and is known to have negative regulatory effects on the immune system (Nakatsukasa et al. 1990, Nelson et al. 1997, Roulot et al. 1999, Sakaguchi et al. 2002). In this study, expression of TGF-β was significantly higher in both the liver (portal space and lobule) and peripheral compartments of the cases compared to the controls. These findings confirm those of other authors (Flisiak et al. 2000, Kinnman et al. 2000, Ray et al. 2003) and provide additional evidence that TGF-β acts as an immune mediator in chronic HCV infection. The negative correlation of S100 DC with peri-portal activity among the cases and its lower expression compared to the controls suggests that there was a lack of HCV recognition by these cells, which, in turn, might have facilitated viral persistence. The mechanisms involved in this down regulation are still unclear and a few hypotheses have been raised. It has been suggested that HCV proteins might modulate T-cell responses by decreasing the stimulatory ability of DC (Sarobe et al. 2003) and that there would be an allostimulatory defect of monocyte-derived DC because these cells would constitute an extra-hepatic reservoir for the virus (Bain et al. 2001). Jinushi et al. (2004) also suggested that the aberrant expression of natural killer (NK) receptors associated with chronic infection might have an impact on the magnitude and direction of DC activation of T-cells. The positive correlation between portal S100 cells and IL-10 in our study reinforces this hypothesis. IL-10 has a modulatory effect on hepatic fibrogenesis by down-regulating the pro-inflammatory response, including the antigen-presenting cells and in a controlled study, IL-10 has also been shown to reduce liver damage (Nelson et al. 2000, 2003). Studies have reported that disease progression is associated with an increased expression of circulating IL-10 (Gramenzi et al. 2005), a correlation with low liver alterations (Piazzolla et al. 2000) and a decreased expression at intrahepatic compartments (Napoli et al. 1996). In our study, the cases had higher serum concentrations of IL-10, but lower intrahepatic expression, than did the respective controls, thus reinforcing the predominance of pro-inflammatory factors. The possible correlations between the duration of infection (n = 41 patients, whose duration of infection could be estimated) or the age of the patient (n = 51) with the immunological parameters was also analysed but did not reveal any biologically meaningful correlations (data not shown). The only possibly relevant finding was the observation that age was positively correlated with portal CD45+RO (p = 0.012, r = 0.35) (as expected), as well as with acinar CD8+ (p = 0.020, r = 0.33), but was negatively correlated with acinar IL-10 (p = 0.024, r = -0.32). Therefore, the duration of the infection or the age of the patient is unlikely to explain our findings. Our study has some limitations. Firstly, it is possible that the observed balance towards a pro-inflammatory type of response was biased by the inclusion of cases with a milder disease; only ~20% presented a more severe classification (F5-F6). For our study, we split the patients into two groups: one group containing the nine patients with severe histology (F5-F6) and the other group containing the 42 patients with mild histology (F1-F4). We then reassessed the correlation analyses of the liver data (cytokine and immunohistochemistry) and blood (cytokines) and we did not find any statistical differences between the two groups in any of the comparisons (data not shown). Also, the data from the 42 patients remained similar to the data from all of the patients (n = 51) (data not shown). Secondly, the cross-sectional nature of the study precludes causal inferences and a prospective assessment of these cases is warranted. However, our results devise several hypotheses for future research. Thirdly, genetic polymorphisms associated with cytokine expression have been shown to play a role in the ability to respond to therapy (Yee et al. 2001, Milton et al. 2005) and were not assessed in our study. Fourth, the function of T lymphocytes was not analysed in this study as it has been in other studies (Moonka et al. 2008). Finally, some comparisons might have been affected by a lack of statistical power, particularly those with a considerable floor effect that was caused by the lower limit of detection for the cytokine assays. In conclusion, we simultaneously assessed pro and anti-inflammatory related cytokines and cell populations in the liver and sera and have shown that (i) the immune response in the samples of predominantly mild disease was associated with several pro-inflammatory cytokines, (ii) CD4+ T-lymphocytes played a major role in orchestrating the immune response and (iii) these events took place primarily at the portal space. Our results suggest that this balance is highly dynamic, particularly in less severe disease and further research is needed to explore the longitudinal aspects of this relationship. REFERENCES
Copyright © 2010 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc10003t4.jpg] [oc10003t1.jpg] [oc10003t3.jpg] [oc10003t2.jpg] [oc10003t5.jpg] [oc10003f2.jpg] [oc10003f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}