 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
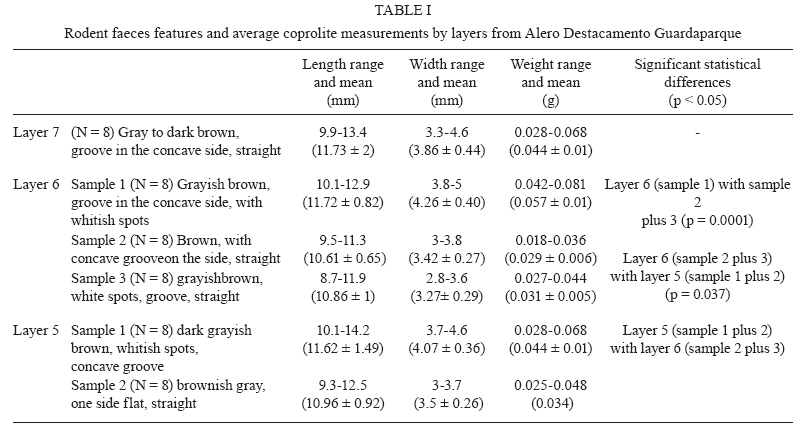
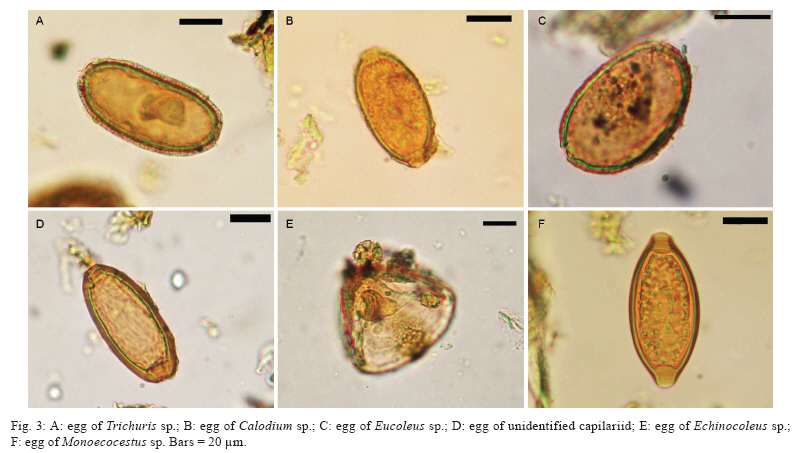
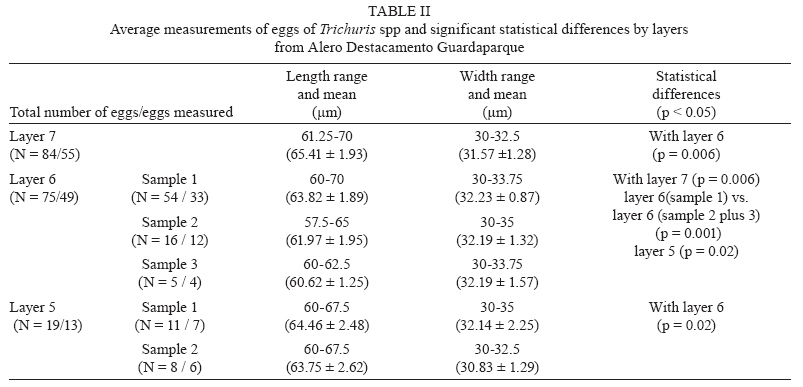
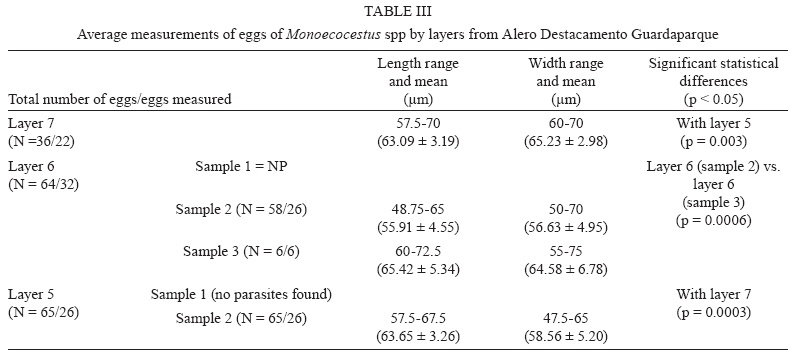
Memórias do Instituto Oswaldo Cruz, Vol. 105, No. 1, 2010, pp. 33-40 ARTICLES Paleoparasitological results for rodent coprolites from Santa Cruz Province, Argentina Norma Haydée SardellaI, IV, +; Martín Horacio FugassaI, IV; Diego Damián RindelII, IV; Rafael Agustín GoñiII, III ILaboratorio
de Paleoparasitología, Universidad Nacional de Mar Del Plata, Funes 3250,
7600 Mar del Plata, Argentina Financial support: FONCyT, Universidad Nacional de Mar del Plata (PICT 06-1520, PICT 07-1576, EXA 438/08), CONICET Received 30 August 2009 Code Number: oc10004 ABSTRACT The aim of this study was to examine the parasite remains present in rodent coprolites collected from the archaeological site Alero Destacamento Guardaparque (ADG) located in the Perito Moreno National Park (Santa Cruz Province, 47º57'S 72º05'W). Forty-eight coprolites were obtained from the layers 7, 6 and 5 of ADG, dated at 6,700 ± 70, 4,900 ± 70 and 3,440 ± 70 years BP, respectively. The faecal samples were processed and examined using paleoparasitological procedures. A total of 582 eggs of parasites were found in 47 coprolites. Samples were positive for eggs of Trichuris sp. (Nematoda: Trichuridae), Calodium sp., Eucoleus sp., Echinocoleus sp. and an unidentified capillariid (Nematoda: Capillariidae) and for eggs of Monoecocestus (Cestoda: Anoplocephalidae). Quantitative differences among layer for both coprolites and parasites were recorded. In this study, the specific filiations of parasites, their zoonotic importance, the rodent identity, on the basis of previous zooarchaeological knowledge, and the environmental conditions during the Holocene in the area are discussed. Key words: paleoparasitology - helminths - rodents - Patagonia - nematodes - anoplocephalids - coprolites Paleoparasitological studies carried out in Argentina are focused on the presence of parasite remains in coprolites, sediments and other materials, from camelids, carnivores, raptor birds, humans and rodents, recovered from Holocenic samples from Patagonia (Fugassa 2006, 2007, Fugassa & Barberena 2006, Fugassa et al. 2006a, b, 2007a, b, 2008a, b, Sardella & Fugassa 2009 a, b). Regarding rodents, their presence in the area seems to have been relevant because of the presence of their faeces, bones and hairs in all archaeological deposits examined at present and also by the finding of rodent remains and helminth eggs in coprolites of rodents and predators (Fugassa et al. 2006b, 2007b, Sardella & Fugassa 2009 a, b). One of the most important archaeological areas under study in Patagonia is the Perito Moreno National Park (PMNP), Santa Cruz Province, Argentina. It exhibits different sites, caves and shelters, with evidences of both human and animal occupations since the Pleistocene - Holocene Transition to the Late Holocene (Aschero et al. 1992, 1993, 2005). The increasing information recovered from other paleoparasitological sites at present, close to Cerro Casa de Piedra (CCP), contributes to enhance the discussion on issues such as the paleoecological biogeography of parasites in the region and the role of zoonoses in the caves and shelters of Patagonia during the Holocene. The aim of the present study was to examine the parasite fauna present in rodent coprolites collected from one of these archaeological sites, the rock shelter Alero Destacamento Guardaparque (ADG). Since 1990 excavations were conducted in the area by archaeologists (Goñi et al. 1994, Goñi 1995, Rindel 2003, 2008), but none paleoparasitological antecedent exists at present. MATERIALS AND METHODS The archaeological site ADG is located in the PMNP (Santa Cruz Province, 47º57'S 72º05'W), close to CCP (Fig. 1). It is a large shelter (250 m length) situated on a low hill oriented towards the southwest. Forty-eight coprolites of rodents obtained from six samples from layers 7 (1 sample, N = 8), 6 (3 samples, N = 24) and 5 (2 samples, N = 16) (Fig. 2), dated at 6,700 ± 70, 4,900 ± 70 and 3,440 ± 70 years BP, respectively, were examined for parasites. Coprolites were inventoried and processed individually as described in Fugassa (2006). The examination consisted of the external observation of faeces (color, texture, inclusions and measurements) according to Chame (2003) and Jouy-Avantin (2003). Faecal macroscopic features and their average measurements are shown in Table I. Each coprolite was fully processed by rehydration in a 0.5% water solution of tris-sodium phosphate in a glass tube for a week; it was then homogenized and allowed to spontaneous sedimentation (Lutz 1919). The material that was sediment into the tube was recovered with a pipette and 10 slides of each sample were observed, with the addition of one drop of glycerine. Eggs of the parasites were measured and photographed at 40X magnification. Statistical analysis was performed using the package Biocistron: Bioestat 5.0 (Hotteling's test). RESULTS Statistical comparisons showed that the coprolites from layers 7 vs. 6 (sample 1), from layer 6 (samples 2 vs. 3) and from layer 5 (sample 1 vs. 2) did not show significant differences (p = 0.09; p = 0.61 and p = 0.06, respectively). Nevertheless, measurements of the faeces from layer 6 (sample 1) vs. 6 (samples 2 plus 3) and 6 (samples 2 plus 3) vs. 5 (samples 1 plus 2) were statistically different (p = 0.0001 and 0.037, respectively) (Table I). These results indicate that the coprolites examined came from different individuals and likely from different host species. Microscopic analysis of the faecal material revealed the presence of plant remains in all samples. The identity of rodents is unknown at present. Of 48 coprolites, 47 were positive for Nematoda and/or Cestoda. In total, 582 eggs were recovered from samples. The parasite contents were as follows: 178 eggs of a species of Trichuris Roederer (Nematoda: Trichuridae), 153 eggs with characteristics attributable to genus Calodium Dujardin, 56 eggs compatible with the genus Eucoleus Dujardin, 29 eggs of an unidentified capillariid, one egg attributable to Echinocoleus sp. López-Neyra (Nematoda: Capillariidae) and 165 eggs of Cestoda (Ano- plocephalidae) with characteristics compatible with the genus Monoecocestus Beddard. For Trichuris sp. (Fig. 3A), eggs were not ornamented and exhibited thick shells and plugs on each end. Average egg measurements from the six samples examined are shown in Table II. Significant statistical differences were found among eggs collected from the three layers and also within each layer. Egg measurements from layer 7 vs. 6 (sample 1) were different (p = 0.006) and also between layer 6 (sample 1) vs. 6 (sample 2 plus 3) (p = 0.001). Among layer 6 (sample 2 plus 3) and 5 (sample 1 plus 2) were not found any difference (p = 0.13 and p = 0.38, respectively), but between layers 6 and 5, the differences were significant (p = 0.02). The identity of the eggs of Trichuris collected from ADG is unknown at present. Eggs of capillariids attributable to the genera Calodium, Eucoleus and Echinocoleus and one unidentified capillariid were found in samples. They were recognized by the typical shape of the ornamented walls. For Calodium sp. (Fig. 3B), large eggs with shallow polar plugs and radial striations were collected. A total of 153 (layer 7, N = 15; layer 6, N = 86 and layer 5, N = 52) eggs was found in all coprolites examined and 97 of them were measured. Average egg measurements from the six samples examined were as follows: layer 7, 62.5-70 (65 ± 2.89) μm long by 33.75-45 (39.11 ± 3.44) μm wide; layer 6: 57.5-75 (65.89 ± 3.43) μm long by 35-45 (39.19 ± 2.45) μm wide and layer 5: 60-62.5 (64.97 ± 3.35) μm long by 35-47.5 (37.71 ± 3) μm wide. No significant differences were confirmed neither among eggs of Calodium collected from layers 7 and 6, nor within 5 (sample 1 plus 2). Nevertheless, egg measurements from layer 5 vs. 6 were different (p = 0.01). Based on the geographic area of the samples and the egg measurements, Calodium from layers 7 and 6 (sample 1) are overlapped and still larger than those reported for Calodium hepaticum (Bancroft) and likely belonged to new species. Eggs attributable to Eucoleus sp. (N = 85) (Fig. 3 C), with irregular striations and short plugs, 50-55 (52.08 ± 2.79) μm long by 22.5-35 (26.61 ± 2.28) μm wide, were collected from the three layers (N = 16, 33 and 7, respectively). Another capillariid eggs (N = 29) (Fig. 3 D) unidentified at present, with pitted surface, 55-67.5 (61.76 ± 3.29) μm long by 32.5-42.5 (37.21 ± 2.61) μm wide were found, 19 of them recovered from layer 6 (sample 1, N = 14, sample 2, N = 4 and sample 3, N = 1) and 10 eggs from layer 5 (N = 7 from sample 1 and N = 3 from sample 2). For both capillariids, no significant differences among layer were registered. Additionally, only one egg compatible with Echinocoleus sp. (65 μm by 31.5 μm) was found from layer 5 (sample 2) (Fig 3E). The surface of the wall of this egg exhibited grooves. A total of 165 eggs of Cestoda (order Cyclophyllidea, family Anoplocephalidae), with features attributable to genus Monoecocestus, was found (Fig. 3F) from the three layers, with exception of sample 1, from both layers 6 and 5. Egg measurements are shown in Table III. Eggs were statistically different among the three layers (p < 0.05). Microscopic remains of rodent hairs, unidentified insect appendages and mites were also discovered in the coprolites. DISCUSSION It was not currently possible to identify the specific biological origin of the rodent coprolites examined. Rindel (2003, 2008) stated that the dominant components of the rodent communities in ADG were Euneomys chinchilloides, Ctenomys sp. and Reithrodon auritus (94%, 93% and 74% from layers 7, 6 and 5, respectively) and in much lesser proportions were present Abrothrix longipilis, Abrothrix xantorhynus, Loxodontomys micropus, Phyllotis xantophyga, Chelemys macronyx, Eligmodontia sp. and Oligoryzomys longicaudatus. Pardiñas and Galliari (2001) noticed that faeces from R. auritus and Euneomys are similar, but those from R. auritus have one end acuminate, unlike those of Euneomys. It is possible that some coprolites examined in the present study belonged to E. chinchilloides or to Ctenomys sp. (without acuminate ends). Redford and Eisenberg (1992) also cited that E. chinchilloides was a dominant species at the end of the Pleistocene, as evidenced by owl pellets contents. The significant differences found in this study among the measurement of faeces collected from the three layers of ADG, could be indicative of the presence of more than one rodent host species in samples. The sites corresponding to the Early Holocene in PMNP are mostly located south of Río Roble. However, a feature that is of interest in ADG, is that despite being in the northern area of Río Roble, the site harbored occupations in early (with contemporary CCP5), as in late stages (Rindel 2003, 2008). One of the assumptions made for the area raises the possible complement functionality between the caves located in the southern sector of PMNP (CCP5 and CCP7) and ADG, for the Early to Medium Holocene. Layer 6 provides chronologies of comparable occupations in CCP5 and CCP7. ADG has been proposed to function as a satellite site of CCP, with limited activities in the rock shelter and numerous activities in the caves (Aschero et al. 1992, 1993, 2005). With respect to the parasites found in samples, it is known that Trichuris spp are parasites of the caecum and colon of mammals of several orders (Anderson 2000). There are many studies on their biology, but little is known about their origin and host's preferences (Robles 2008). In America there were identified 21 species of Trichuris in nine rodent families, with five species found in Argentina, Trichuris dolichotis Morini, Boero and Rodriguez, parasite of Dolichotis patagonum (Zimmermann) (Morini et al. 1955), Trichuris laevitestis Suriano et Navone, parasite of Scapteromys aquaticus Thomas and Akodon azarae (Fischer) (Suriano & Navone 1994, Robles & Navone 2006); Trichuris bursacaudata Suriano et Navone, parasite of Ctenomys talarum Thomas (Suriano & Navone 1994); Trichuris pampeana Suriano et Navone parasitic on Ctenomys azarae Thomas (Suriano & Navone 1994, Rossin & Malizia 2005) and Trichuris pardinasi Robles, Navone et Notarnicola parasite of Phyllotis xanthopyga (Waterhouse) (Robles et al. 2006, Robles 2008). Reports of Trichuris spp in South American rodents from ancient material are those of Ferreira et al. (1991), Araújo et al. (1993), Fugassa (2006) and Sardella and Fugassa (2009a, b), among others. Because of the overlap in the egg dimensions for Trichuris spp infecting rodents, not only in Argentina, but also in Chile, as is the case for Trichuris bradleyi, Trichuris chilensis, Trichuris fulvi and Trichuris robusti (Babero et al. 1975, 1976, Babero & Murúa 1987, 1990), the parasites found in ADG were, therefore, assigned to more than one species of Trichuris. In this sense, there were differences in the archaeofaunistic composition among the layers in ADG, showing different percentages in the rodent composition of communities throughout the studied period (Rindel 2003, 2008). Based on the geographic area of the samples and the egg measurements, eggs of Trichuris from layers 7 and 6 (sample 1) are within the range of those reported for T. bursacaudata (Suriano & Navone 1994). Trichuris from layer 6 (sample 2 plus 3) corresponds to T. bradleyi (Babero et al. 1975, 1976, Babero & Murúa 1987, 1990) and Trichuris from layer 5 is within the range of Trichuris sp. (layer 13 from CCP7, dated at 7,920 ± 130 yr BP) (Sardella & Fugassa 2009b), or they also could be new species, not yet described for the studied area. Some authors argue that Trichuris is not present in extreme latitudes, low temperatures being the main limiting factor (Nolf 1932) and that the areas around the tropics (warm and humid) would offer more favorable conditions for the development and survival of various parasites (Bundy & Cooper 1989). Nevertheless, the presence of Trichuris in ADG and from all the archaeological sites examined from Patagonia at present, help to doubt those assumptions. Classification of Capillariidae is one of the most complexes among nematodes (Anderson 2000). The family is composed of 22 genera and 300 species (Anderson 2000, Moravec 2000, 2001). In America, 16 species of Capillaridae are known as parasites of 10 families of mammals, four of them are parasites of rodents of the families Cricetidae, Muridae, Cavidae and Sciuridae (Robles 2008). In Argentina, Eucoleus gastricus (Baylis) was reported as the parasite of Rattus norvegicus (Berkenhout) (López-Neyra 1947, in Robles 2008). Robles (2008) consigned that in sigmodontine rodents, three species of Capillariidae with distribution in North America, C. hepaticum, E. gastricus and Aonchotheca forresteri (Kinsella et Pence) were recognized. Robles (2008) found Eucoleus sp. and Echinocoleus sp. in Akodontini rodents from Cuenca del Plata, Buenos Aires Province. Paleoparasitological records of capillariids indicate their ubiquitous presence in several samples and from different archaeological sites (Fugassa 2006, 2007, Fugassa & Barberena 2006, Fugassa et al. 2006a, b, 2007a, b, 2008a, b, Sardella & Fugassa 2009 a, b). The presence of eggs attributable to Calodium sp., Eucoleus sp. and Echinocoleus sp. adds the rock shelter ADG to the previous records. In relation to Calodium, egg measurements are in line and still larger than the eggs of C. hepaticum. Moreover, the life-cycle of C. hepaticum assumes the appearance of little or no egg in faeces. There are major limitations in identifying specimens of Capillaria sensu lato to species level. Zhu et al. (2000), using mitochondrial DNA analysis within and among species of Capillaria s. l. from Australian marsupials and rodents, cited that, although highly variable in their host and tissue specificity, they may exhibit the greatest degree of specificity at the level of host genus. This argument, together with the differences in measurements found, allow to state that the eggs of capillariids examined from ADG could belong to new species, not yet described for the area. The genus Eucoleus is currently distributed in America, Europe, Oceania and Asia and includes 30 species parasitizing the respiratory organs, oral cavity, tongue, oesophagus and stomach of birds and mammals (Moravec 2001). Robles (2008) found 2 species of Eucoleus from the stomach of two species of cricetids collected from North of Argentina. Respect to the paleoparasitological records of this parasite genus, Sardella and Fugassa (2009b) found eggs in coprolites of Ctenomys sp. from CCP7 and in this study, the presence of Eucoleus sp. was confirmed throughout all the Holocenic period of time considered, present in the three layers. The genus Echinocoleus is currently distributed in America, Asia and Oceania and comprises six species parasitizing the intestine of birds and mammals (Moravec 2001). The first communication of the presence of this genus in Argentina was that of Robles (2008), who found Echinocoleus sp. from one cricetid from Buenos Aires province. The finding of only one egg of this nematode in the present study was probably due to a low parasitic load or taphonomic processes. Anoplocephalidae cestodes are represented by 25 genera in small mammals (Wickström et al. 2005). Intermediate hosts are oribatid mites ingested by their herbivorous definitive hosts (Beveridge 1994). Rego (1961) reviewed the genus Monoecocestus, recording several species from rodents living in Brazil, Paraguay, Perú and Chile. In Argentina, Sutton (1973) described the species Monoecocestus myopotami, in the otter Myocastor coypus. The only previous paleoparasitological record of this genus in rodents was from coprolites from the historical site Alero Mazquiarán (Sardella & Fugassa 2009a). Eggs of Monoecocestus found in the present study from ADG probably belonged to different species, because they exhibited significant differences among layers. It should be noted that all the parasitic representatives of the genera found in this study are currently known as zoonotic. Perkins et al. (2005) stated that rodents represent one of the most important sources of zoonoses for mammals and that the increasing density of rodents forced their dispersion to occur and brought them into closer contact with humans. Species of Trichuris are included among the zoonotic geohelminths (Wong & Bundy 1990), since infection involves ingestion of contaminated soils. Bundy & Cooper (1989) stated that gastrointestinal helminthosis is estimated to be the most common of all parasitic infections, with more than a quarter of the world's population infected with four human nematode species, Trichuris trichiura Linnaeus occupying the second order of importance, after Ascaris lumbricoides Linnaeus. The zoonotic potential of Trichuris was also documented for Trichuris vulpis, parasite of dog worldwide (Singh et al. 1993, Dunn et al. 2002). It is probable that Trichuris spp of Patagonic rodents in ADG could be infectious for humans, because during the Holocene, humans and animals shared caves and shelters. Capillariidae exhibit both direct and indirect cycles, a vertebrate being the definitive host. In the case of heteroxenic cycles, the intermediate host ingests the eggs and the infective larvae develop in it (Anderson 2000, Morand et al. 2006). The life-cycle of C. hepaticum requires rodent death, ingestion and digestion of infected liver and then deposition of unembryonated eggs in the environment, which will allow them to survive and embryonate; mice are thought to become infected through ingestion of embryonated eggs (Singleton et al. 1991). Reports of human infections with C. hepaticum in different parts of the world could indicate that infection is more common in humans than supposed and the majority of proven infections have ended fatally (Farhang-Azad & Schlitter 1978). It is unlikely that Calodium spp found in rodent coprolites from ADG was C. hepaticum. However, it seems likely that other capillariid parasites of rodents, not only C. hepaticum, may have potentially caused disease, because of the close contact of humans with rodents in the Holocenic caves and shelters. The life cycle of Eucoleus aerophilus is not well known at present, but it appears to involve direct transmission, as well as indirect transmission through earthworms, to canids, felids and foxes (Radman et al. 1986, Burguess et al. 2008). In relation to the hazards to humans, Eucoleus sp. has been reported as a human parasite in seven cases in Russia, one in Morocco, one in Iran and one in Ukraine (Beaver et al. 1984, in Bowman et al. 2002). The most common signs were acute bronchitis and broncheolitis, usually with asthma and cough. Despite eggs found in ADG could not be identified at specific level, the role of Eucoleus sp. as possible causative of illness to humans could not be discarded. Knowledge of Echinocoleus life cycle and pathogeny is limited, because considerations on its zoonotic importance are inexistent in bibliography at present. Moravec (2001) stated that, of 22 present valid genera of capillariids, the life cycle has been studied in members of only 10 genera and that there are not data in this sense concerning the genus Echinocoleus. Anoplocephalids can cause human disease when humans eat mites present in the soil (Denegri et al. 1998). Humans living in ADG were probably exposed to illness by Monoecocestus spp during all the period of time considered. The knowledge of the climatic conditions prevalent in the area during the Early to Medium Holocene has conducted studies on the environmental conditions during the period in Patagonia (Stine & Stine 1990). The sequence in the Patagonian climate was similar throughout the Holocene, with a trend toward aridity since the Middle Holocene. However, the information generated by different lines of research on a regional scale, suggests substantial fluctuations in climate and landscape to be considered in the reconstruction of settlement and mobility strategies of hunter-gatherer populations. One source of information available was studied from the Lago Cardiel, 110 km SE of ADG. Stine and Stine (1990) indicate significant variations in the level of the lake in the Holocene, alternating dry periods with episodes of elevation of the moisture. These authors identified peaks of the lake height between 10,000-7,000 years BP, then between 5,500-4,500 years BP. Subsequently, these levels were never reached again, alternating periods of very low humidity with some increases. Another kind of paleoenvironmental information proceeded from the pollen studies carried out in the area, showing the alternation between the shrubland and herbaceous steppe, as well as the variability registered in the extent of Nothofagus forest, over the past 7,000 years (Mancini et al. 2002). The analysis suggests that in environments such as the mosaic of the study area, plant communities react sensitively and rapidly to extreme climatic oscillations. The study presented here suggests that, sites of habitual occupation by Patagonian hunters such as caves and shelters, could have acted as potential environments for transmission of various types of parasites, as those identified here. Under these conditions, the study of lines of paleoparasitological evidence is presented as an important source of information about the relationships that the Patagonian hunter-gatherer groups established with the environment, information that can be combined in future with other disciplines, such as the zooarchaeology and bioarchaelogy, in order to obtain a better picture of the lives and health of past human groups. REFERENCES
Copyright © 2010 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc10004f3.jpg] [oc10004f1.jpg] [oc10004f2.jpg] [oc10004t1.jpg] [oc10004t2.jpg] [oc10004t3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}