 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
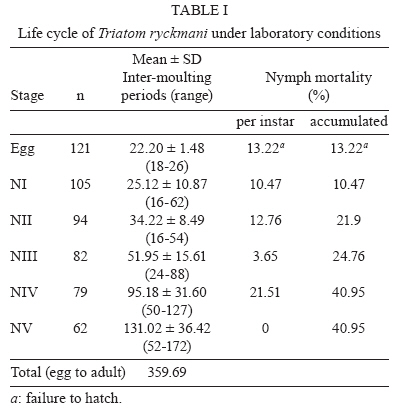
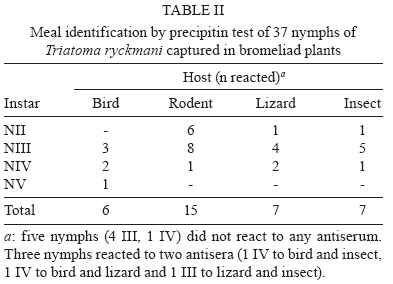
Memórias do Instituto Oswaldo Cruz, Vol. 105, No. 1, 2010, pp. 99-102 ARTICLES Life cycle of Triatoma ryckmani (Hemiptera: Reduviidae) in the laboratory, feeding patterns in nature and experimental infection with Trypanosoma cruzi Rodrigo ZeledónI, +; Marlen CorderoI; Ricardo MarroquínII; Elias Seixas LorosaIII ILaboratorio
de Zoonosis, Escuela de Medicina Veterinaria, Universidad Nacional, Heredia,
Costa Rica Received 28 October 2009 Code Number: oc10015 ABSTRACT A cohort initiated with 121 eggs, yielding 105 first instar nymphs (eclosion rate: 86.78%), allowed us to observe the entire life cycle of Triatoma ryckmani under laboratory conditions (24ºC and 62% relative humidity), by feeding them on anesthetized hamsters. It was possible to obtain 62 adults and the cycle from egg to adult took a mean of 359.69 days with a range of 176-529 days (mortality rate of nymphs: 40.95%). Mean life span of adults was of 81 days for females and 148 days for males. The developmental periods of 4th and 5th nymphs were longer than those of the other instars. This suggests that young siblings have a better chance of taking a hemolymph meal from older ones, in order to survive during fasting periods during prolonged absences of vertebrate hosts from natural ecotopes. The stomach contents of 37 insects showed blood from rodents (15 cases), lizards (7 cases), birds (6 cases) and insect hemolymph (7 cases). Out of 10 insects fed by xenodiagnosis on a Trypanosoma cruzi infected mouse, all but one became infected with the parasite. Key words: Triatoma ryckmani - life cycle - food preference - T. cruzi Triatoma ryckmani was first described from a single female specimen found in a military barrack in Valle Department, Honduras, Central America (Zeledón & Ponce 1972). Later on, three more female specimens were found in airplanes in Miami, United States, one from Honduras and two from Guatemala, the latter together with bromeliad plants in air freight (Lent & Wygodzynsky 1979). The first male specimen was found near the town of Santa Cruz, Guanacaste Province of Costa Rica (Sherlock & Morera 1988). Carcavallo et al. (1996) did an external morphological study, with scanning electron microscopy, of another male specimen, collected in Santa Rosa, Guanacaste Province, Costa Rica. More recently, the natural habitats of T. ryckmani were discovered in a semiarid region of El Progreso Department, Guatemala, in cacti (Stenocercus eichlamii) and epiphytic bromeliads (Tillandsia xerographica). In the first instance, all instars of the bug were found in the dry portions of the cactus (Marroquín et al. 2004a, Monroy et al. 2004); in the second situation, colonies of the bugs were also found within the leaves of the bromeliads growing on trees, including the cacti (Marroquín et al. 2004b). The species has also been found in at least four Departments of Nicaragua in small numbers in houses, under intra and peridomiciliary conditions (Marín et al. 2006). Here we describe, for the first time, the entire life cycle of T. ryckmani under laboratory conditions and the blood patterns of the different instars from specimens collected under natural conditions in Guatemala. MATERIALS AND METHODS A colony was started from specimens captured in June 2005 in bromeliads from a semidesertic area of the Department El Progreso, Guatemala, 89 km from the capital. The climate is hot and dry with a mean maxima temperature of 34.3°C (2003-2008) and a mean annual rain fall of 762.2 mm for the same period. A cohort was initiated with 121 eggs laid, during a period of six weeks, by one female captured as a 5th instar nymph. The eggs, obtained during different days, were separated in nine groups of 8-18 eggs, by date of oviposition, in 10 cm diameter plastic Petri dishes with filter paper on the bottom. They were incubated at a constant temperature with relatively little variation during the experiment (mean ± SD: 26.0 ± 0.37) and 62.0 ± 3.32 relative humidity. Each group was fed on anesthetized hamster (0.25 mL of a mixture of Ketamine, 3 parts and xylazine, 1 part) by xenodiagnosis in a small box, during 40-60 minutes, every two weeks and evidence of ecdyses was monitored daily. First instar nymphs were fed at 4-5 days after hatching. Up to the 4th instar, the nymphs required from 2-4 meals for molting; 4th and 5th instars required 5-7 meals and 9-12, respectively, even though they usually fed readily to repletion. The sources of blood meals of 37 insects, obtained from T. xerographica, were collected individually from the stomach in separate filter papers for each one and after eluded they were identified by the capillary tube precipitin test with antisera against 15 different hosts (human, fowl, dog, cat, horse, goat, cow, pig, sheep, rodent, opossum, armadillo, toad, lizard and insect hemolymph), following the procedure of Lorosa et al. (1998). In addition, 10 individuals (2 adult males, 3 4th instar nymphs and 5th instar nymphs) were fed by xenodiagnosis on a mouse infected with a Costa Rican human strain of Trypanosoma cruzi (GA strain) and observed periodically after one week for a period of 30 days. RESULTS Details of the period required for the eggs to hatch and for each nymphal instar to molt until adulthood are presented in Table I. It can be observed that the life cycle of T. ryckmani from 1st up to the 4th instar nymph is rather fast (111.29 as a mean), as compared with the molting periods of the 4th and 5th nymph stages together (226.20 days). The entire life cycle from egg to adult took about a year (359.69 days as a mean of means) with extremes between 176-529 days. The figures for 5th instar nymphs producing males were 118.29 ± 28.93 days as a mean, with extremes between 67-172 days and a total life cycle of 346.96 days as a mean (31 individuals); and the same periods for females were 143.87 ± 38.96 days with extremes of 52-236 and a total of 392.54 days, also for 31 specimens. The eclosion rate was high (86.78%) and total nymph mortality rate reached 40.95% during the entire cycle. No mortality was observed in the 5th instar nymphs and the lowest rate was observed in 3rd instar nymphs (3.65%). The highest mortality occurred at the 4th stage (21.51%). The life span of adults was 81.05 ± 30.21 days for females with extremes of 78-135 and 148.13 ± 38.24 days with extremes of 33-159 for males. The meals found in the stomachs of the insects reacted only against four antisera: fowl, rodent, lizard and insect hemolymph. Out of 37 samples tested, all of different instar nymphs, five did not react against any of the antisera, 29 reacted to at least one of them and three reacted to two hosts at the same time (Table II). The most common host identified was rodent. It is important to notice that seven of the reacting meals revealed that these nymphs had fed on another insect. Of the 10 insects used in xenodiagnosis on a mouse infected with T. cruzi (2 adults and 8 nymphs) all but one 4th instar nymph became infected, showing metacyclic trypomastigotes in their dejections. DISCUSSION The different methodologies used to study the duration of the life cycles of triatomine species under laboratory conditions make it difficult to compare the information gathered. Nevertheless, it is now clear that several intrinsic factors characteristic of a particular species, environmental conditions (mainly temperature and humidity), blood sources, frequency of feeding and other factors can influence the time for the bugs to reach adulthood (Guarnieri et al. 2000, Emmanuelle-Machado et al. 2002, Damborsky et al. 2005). We also know that, in general, there is an increase in time between molts as the insect grows. Some species have the potential to produce more than one generation a year while others need one or two years or even more to complete the cycle and, in some cases, a full blood meal is generally enough to allow molting (Zeledón et al. 1970, Galvão et al. 1995, Rocha et al. 2004, Luitgards-Moura et al. 2005, Villacis et al. 2008). Some food sources may be insufficient for the completion of the entire cycle of some species, but not necessarily in others (Guarnieri et al. 2000, Rocha et al. 2001, Emmanuelle-Machado et al. 2002). We have to keep in mind that it is almost impossible to reproduce in the laboratory the conditions under which the insects live in nature, especially for those which use particular ecotopes with a suitable microclimate. The delayed time required for the 4th and 5th nymph instars of T. ryckmani to reach the molting periods deserves special comment. The mean time periods of these instars added and divided by the other three together, yields a rate that is twice that of the first three instars (2.0). If we calculate the same rate in other species of triatomines (sum of developmental periods of 4th and 5th instar nymphs/sum of developmental periods of 1st, 2nd and 3rd instar nymphs) we find that this is not the case for them. The mean for 15 other species (Triatoma flavida, Triatoma nitida, Triatoma maculata, Triatoma klugi, Triatoma gerstaeckeri, Triatoma lecticularia, Triatoma protracta, Triatoma mexicana, Triatoma dimidiata, Meccus pallidipennis, Meccus longipennis, Rhodnius neglectus, Rhodnius brethesi, Rhodnius ecuadoriencis and Panstrongylus rufotuberculatus) is 1.2 with a range of 0.7-1.8 (Zeledón et al. 1970, Galvão et al. 1995, Martínez-Ibarra et al. 1999, 2003, 2007, 2008, Cabello & Lizano 2001, Rocha et al. 2001, 2004, Emmanuelle-Machado et al. 2002, Wolf et al. 2004, Luitgards-Moura et al. 2005, Villacis et al. 2008). This phenomenon in T. ryckmani suggests long periods of fasting by the nymphs, which is probably related to prolonged absences of a vertebrate host from the natural biotopes. One of us (R Marroquín, unpublished observations) has observed a rodent (Reithrodontomys sp.), a bird (Columbina sp.) and a lizard (Ctenosaura palearis), visiting the natural habitats. Under these conditions, larger nymphs, which might endure longer periods of fasting, might serve as a food source for younger nymphs by cannibalism. An evolutionary adaptation process probably allows larger nymphs to prolong their nymphal periods, in order to guarantee the survival of the smaller ones. Cannibalism is a well known phenomenon in triatomines (Zeledón & Rabinovich 1981) and probably plays an important role in the survival of the nymphs of several species. Hays (1965) observed cannibalism in Triatoma sanguisuga and was able to make a 1st instar nymph to reach the 4th stage fed only on hemolymph and believes that the process is probably useful in the survival of early-instar nymphs. Cannibalism might occur not only among the nymphs of the same species, but sometime with other types of insects or invertebrates that are readily available (Carvalho-Pinto et al. 2000). The fact that we found insect hemolymph in the stomach contents of seven out of 37 nymphs (1 of 2nd instar, 5 of 3rd and 1 of 4th) seems to reinforce this hypothesis. Also, Lorosa et al. (2000) were able to rear Triatoma rubrovaria and Triatoma circunmaculata in the laboratory by feeding them on large roaches of the genus Hormetica. Despite observing high mortality during the life cycle, particularly in the 4th instar, it was possible to obtain some adults of both species by this mechanism. In spite of the fact that T. ryckmani has not been found infected with T. cruzi under natural conditions (Marroquín et al. 2004a, Monroy et al. 2004, Marín et al. 2006), the species is able to colonize chicken coops, sometimes in elevated numbers, up to 369 in a single coop (Nakagawa et al. 2005). It also has been found colonizing houses, in smaller quantities, and adults are also found visiting houses in domestic and peridomestic environments, both in Nicaragua and Guatemala (Monroy et al. 2004, Nakagawa et al. 2005, Marín et al. 2006). Now that we know that this species becomes easily infected with T. cruzi in the laboratory, its natural infection could be predicted and its epidemiological role in the transmission of Chagas disease in Central American countries needs to be assessed. ACKNOWLEDGEMENTS To Dr. Paul Hanson (School of Biology, University of Costa Rica), for his revision of the manuscript, and to Mr. Heri-berto Gutiérrez, for valuable technical help. REFERENCES
Copyright © 2010 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc10015t2.jpg] [oc10015t1.jpg] |
| |||||||||

{kind=link}
{kind=link}