 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
SHORT COMMUNICATION Occurrence and composition of class 1 and class 2 integrons in clinical and environmental O1 and non-O1/non-O139 Vibrio cholerae strains from the Brazilian Amazon Lena Lillian Canto de SáI; Érica Lourenço da FonsecaII; Maíra PellegriniII; Fernanda FreitasII; Edvaldo Carlos Brito LoureiroIII; Ana Carolina Paulo VicenteII, + ILaboratório de Microbiologia Ambiental, Seção de Meio Ambiente Received
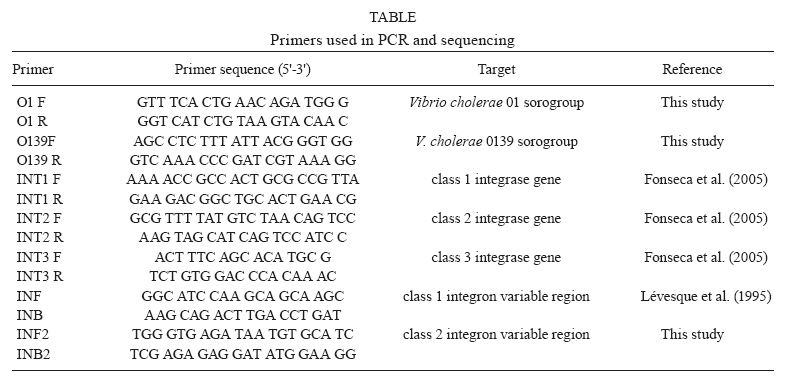
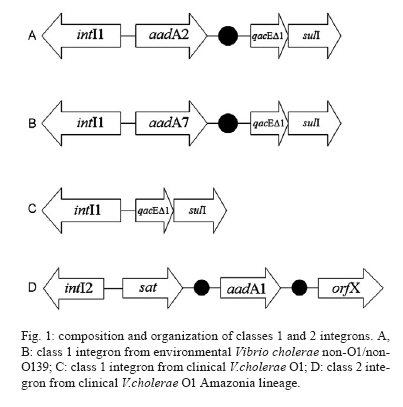
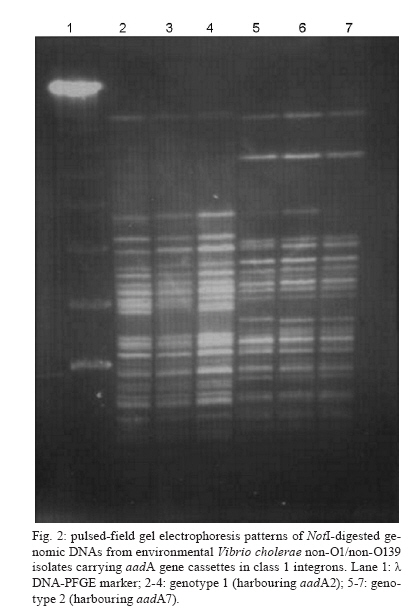
19 August 2009 Financial support: IOC, IEC/SVS/MS, FAPERJ, CNPq Code Number: oc10040 ABSTRACT This study identified and characterised class 1 and 2 integrons in clinical and environmental Vibrio cholerae O1 and non-O1/non-O139 strains isolated from the Brazilian Amazon. The aadA2 and aadA7 gene cassettes were found in class 1 integrons in two genotypes of environmental V. cholerae non-O1/non-O139. Empty integrons were found in strains from the Brazilian cholera epidemic. A class 2 integron was detected in one strain from the V. cholerae Amazonia lineage harbouring sat1 and aadA1 genes. All isolates were resistant to aminoglycosides, indicating aadA functionality. These findings suggest that environmental bacteria act as cassette reservoirs that favour the emergence of resistant pathogens. Key words: Vibrio cholerae - class 2 integron - Amazon The integrase (IntI) is the signature of an integron. Three classes of integrons (classes 1, 2 and 3) have been described as resistant integrons (RIs), which can contain several antibiotic resistance genes in tandem. Class 1 is recognised as the most widespread integron found in clinical bacterial isolates. This element is characterised by a 5' conserved segment (CS), which includes the intI gene, the recombination site (attI) and a promoter region (Pc) and a 3' CS, which usually includes the qacEδ1 and sulI genes. Between these CSs, there is a variable region where gene cassettes can be inserted and expressed under Pc control (Stokes & Hall 1989, Recchia & Hall 1995, Rowe-Magnus & Mazel 2002). Class 2 integrons are not as widespread among bacteria as class 1 integrons, even though class 2 integrons are associated with a mobile DNA element, the Tn7 transposon. Class 2 integrons harbour an inactive intI due to a premature stop codon, which could explain the reduced variety and number of cassettes, frequently only sat and aadA1. The mobilisation of gene cassettes in class 2 integrons can occur due to in trans activity of other IntIs present in the same bacterial cell (Hansson et al. 2002). Most of class 1, 2 and 3 integrons were recovered from bacteria strains isolated in clinical settings; however, there is evidence that gene cassettes are transferred between environmental, commensal and pathogenic bacteria harbouring distinct integron classes (Michael et al. 2008, Gillings et al. 2009). In this context, class 1 integrons retain the gene cassettes that confer adaptive advantages to environmental pressures (Hardwick et al. 2008). In this work, we identified and characterised, for the first time, class 1 and 2 integrons in clinical and environmental Vibrio cholerae O1 and non-O1/non-O139 isolates from the Brazilian Amazon. This study was performed with 126 V. cholerae non-O1/non-O139 isolates obtained from river water and sewage from the Amazon Region between 1977-2006 and with 60 clinical isolates recovered during the South American cholera epidemic (1991-1995). All isolates were cultured on TCBS agar plates and biochemically identified using API 20E. Serogroups were defined by PCR using specific primers for serogroups O1 and O139 (Table) and also by agglutination test. Susceptibility tests performed with VITEK® cards (bioMérieux, Marcy l'Étoíle, France) demonstrated that the majority of isolates were sensitive to all antibiotics. In order to verify the presence of RIs (classes 1, 2 and 3) in V. cholerae strains, PCR reactions were performed using primers targeting the intI genes, the variable regions from class 1 and 2 integrons and the 3' CS from class 1 integrons (Table). The PCR reactions were performed as described elsewhere (Fonseca et al. 2005). The amplicons were purified using Wizard SV Gel and PCR Clean-UP System kits (Promega) and both strands were directly sequenced using Big Dye Terminator v 3.1 Cycle Sequencing Ready Reaction Kit in a 3100 Automated DNA Sequencer (Applied Biosystems). Nucleotide sequences were compared to those available in the GenBank database, which can be found at the website http://www.ncbi.nlm.nih.gov. The genetic relatedness of the strains that carried the integrons was determined by DNA macrorestriction using NotI followed by pulsed-field gel electrophoresis (PFGE). Band profiles were compared visually and interpreted according to the criteria described by Tenover et al. (1995); for example, an isolate was considered to belong to a genotype if its macrorestriction profile presented at most three different bands. The intI1 gene was detected in 7/126 (5.5%) environmental non-O1/non-O139 strains. A molecular survey of integrons in V. cholerae non-O1/non-O139 strains in an aquatic environment of Tehran (Iran) found a prevalence of 5.4% (Bakhshi et al. 2009), almost the same rate as that observed in our study. The sequence analysis of the integron variable regions revealed the presence of two alleles of the aadA gene, aadA2 and aadA7 (GenBank accessions DQ196321 and DQ196322, respectively) (Fig. 1), which confer resistance to aminoglycosides. Each distinct allele was found in V. cholerae non-O1/non-O139 strains from two genotypes as determined by PFGE analysis (Fig. 2). Strains from genotype 1 were isolated from sewage water in 1977 and carried the aadA2 gene. Interestingly, strains from genotype 2, harbouring the aadA7 gene, had persisted in sewage from 1978-1981. In a survey of 3,000 Gram-negative bacteria from an estuarine environment in England performed over a two-month period, the prevalence of class 1 integrons was 3.6% and aadA1 was the most frequent gene cassette found (Rosser & Young 1999). A total of 20/60 (33.3%) clinical V. cholerae O1 strains from the cholera epidemic in the Amazon (1991-1995) harboured the intI1 gene and intI2 was found in only one isolate. No intI3 gene was detected. These clinical strains belonged to two distinct V. cholerae O1 lineages. All strains that contained the intI1 gene belonged to the South American cholera El Tor epidemic genotype (Vicente & Coelho 2005). Analysis of the variable region in these strains revealed no gene cassettes, indicating the occurrence of empty class 1 integrons (Fig. 1). These data have been observed by others (Rosser & Young 1999). However, the qacEδ1 and sulI genes, which define the 3'CS, were present. This result contrasts with a previous work that was unable to show class 1 integrons in V. cholerae O1 strains from the Brazilian cholera epidemic (Campos et al. 2004). A class 2 integron was identified in only one V. cholerae O1 clinical strain that belonged to the Amazonia lineage (Coelho et al. 1995). Sequence analysis of the 3.3 kb amplicon, obtained with the primers INT2 F and INB2, revealed the presence of the intI2-sat-aadA1-orfX gene array, corresponding to an inactive IntI, a streptothricin acetyltransferase, a streptomycin 3''adenyltransferase and a hypothetical protein (GenBank accession DQ196320). Interestingly, the dihydrofolate reductase type I (dfrA1) gene, prevalent in most class 2 integrons, was missing in the cassette array identified here. The strains that carried class 1 integrons, with aadA alleles and the isolate harbouring a class 2 integron were resistant to streptomycin and spectinomycin, which indicates the functionality of these aminoglycoside resistance genes. Class 1 integrons carrying distinct cassette arrays have been detected in V. cholerae O1 and non-O1/non-O139 strains in Europe and Asia, indicating the heterogeneity in organisation and/or composition of the variable region (Lévesque et al. 1995, Dalsgaard et al. 2000). Here, clinical V. cholerae O1 strains from the Brazilian cholera epidemic were shown to carry empty class 1 integrons. Conversely, V. cholerae non-O1/non-O139 environmental isolates recovered from a period before the cholera epidemic carried class 1 integrons harbouring alleles of the aadA gene. The aadA2 allele is widespread among bacteria species around the world. Interestingly, aadA2 was in the first position in a class 1 integron cassette array found in a V. cholerae O1 clinical strain isolated from the Amazon Basin in 1998. In this array, the newly described qnrVC1 gene cassette was also identified (Fonseca et al. 2008). These findings provide evidence of environmental bacteria acting as reservoirs for particular gene cassettes that favour the emergence of multi-resistant pathogenic isolates. Of note was the identification of a V. cholerae strain persisting in the environment for four years and harbouring class 1 integrons containing the aadA7 gene. Such persistence indicates that this strain not only shows a fitness for this environment, but may also horizontally transfer the aadA7 gene cassette to other bacteria carrying integrons. Thus far, the presence of class 2 integrons in V. cholerae was shown in two distinct isolates of V. cholerae non-O1/ non-O139, one from India (2003) and the other from Bangladesh (2005) (Ahmed et al. 2006) and in V. cholerae 01 isolates from an outbreak in Ghana (Opintan et al. 2008). The three strains were carrying typical class 2 integrons with the same resistance gene cassette array, dfrA1/sat1/aadA1. The presence of the rare class 2 integron in only one strain of the V. cholerae O1 Amazonia lineage, isolated in 1991, is evidence of the dynamic process of integron mobilisation and gene cassette loss and acquisition, as well gene spreading among other bacterial species. Our results highlight the importance of considering environmental bacteria in surveillance programs that focus on detection of genetic elements related to lateral gene transfer and antibiotic resistance and contribute to our understanding of the emergence of resistant bacteria in clinical settings. ACKNOWLEDGEMENTS To PDTIS sequencing platform from Fundação Oswaldo Cruz. REFERENCES
Copyright © 2010 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc10040f1.jpg] [oc10040t1.jpg] [oc10040f2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}