 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
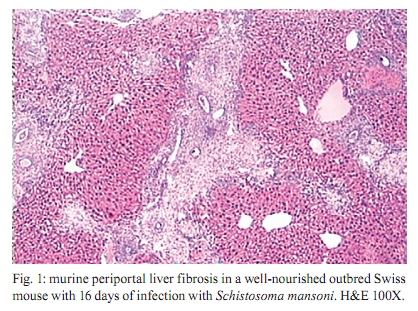
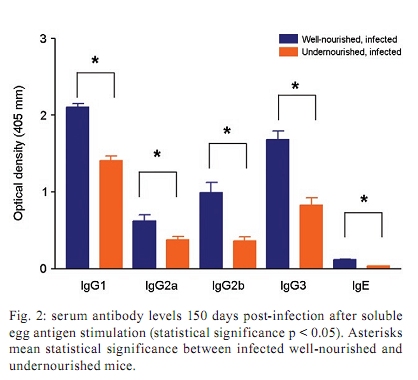
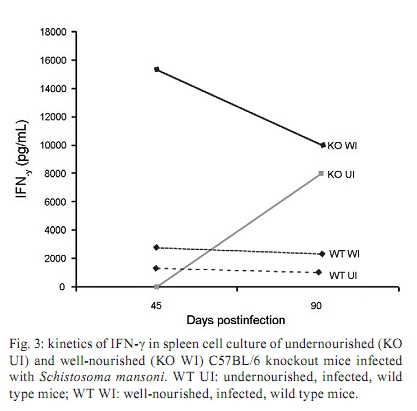
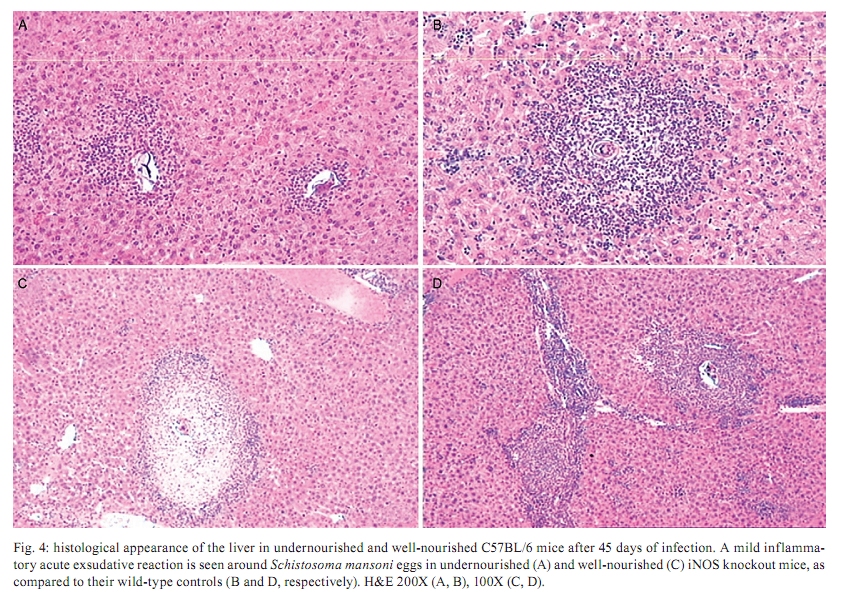
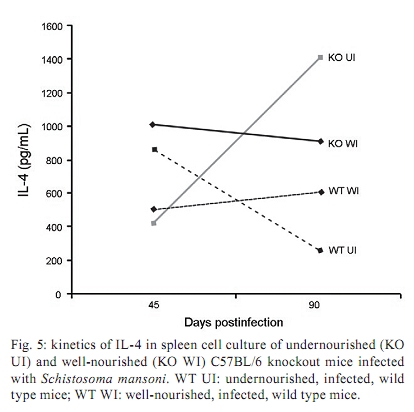
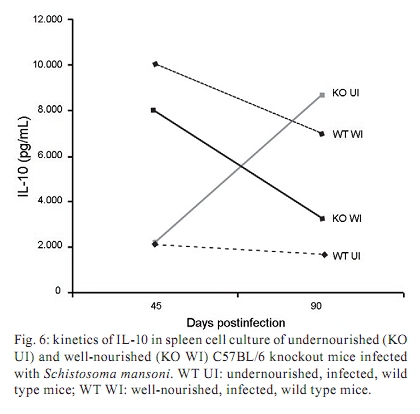
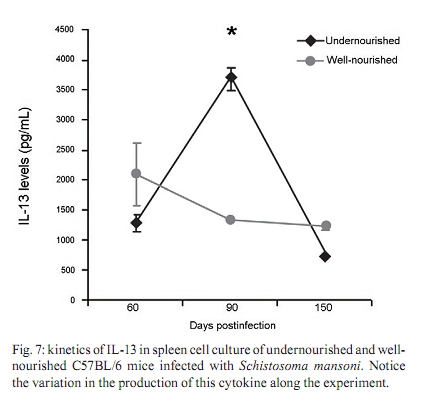
Memórias do Instituto Oswaldo Cruz, Vol. 105, No. 4, 2010, pp. 359-366 Manson's schistosomiasis in the undernourished mouse: some recent findings Eridan M Coutinho+; Sheilla A de Oliveira; Andréia F de Barros; Fabiana L Silva; Renata P Ramos Laboratório de Imunopatologia e Biologia Molecular, Centro de Pesquisas Aggeu Magalhães-Fiocruz, CP 7472, 50670-420 Recife, PE, Brasil + Corresponding author: eridan@cpqam.fiocruz.br Received 9 January 2009 Code Number: oc10063 ABSTRACT This paper deals with current knowledge of the interrelationships between Schistosoma infection and malnutrition. It emphasizes the relevance of these investigations in the face of dynamic and evolving changes occurring in population diets and changes in the epidemiological patterns of schistosomiasis in endemic countries. The paper further discusses the basis for continuing the studies on this subject and the reasons why it represents a misunderstood association. This review also focuses on the cellular and humoral immune responses in the undernourished mouse model infected with Schistosoma mansoni, with updated information on the immune response in wild-type and iNOS knockout mice concerning soluble egg antigen specific antibodies and kinetics of IFN- γ , IL-4, IL-10 and IL-13 cytokines, in the chronic phase of Manson's schistosomiasis. There is indication that schistosome-infected undernourished mice are able to develop a humoral immune response, but antibody titres are much lower than in the control animals. Cytokine production (IFN- γ , IL-4, IL-10) is lower in the undernourished mice, but as infection progresses to the chronic phase its kinetics run an antagonistic course when compared to that of well-nourished animals. Marked variation in the secretion of IL-13 (a fibrogenic cytokine) could explain why undernourished mice do not develop liver "pipe-stem" fibrosis described in previous papers on well-nourished animals. Key words: Schistosoma mansoni - undernutrition - immunopathology Based on the literature on the interaction between malnutrition and schistosome-infection, it can be concluded that it is still necessary to get a deeper insight into the subject, considering incomplete information and sometimes conflicting results reported in some publications. The purpose of this paper is to stress the relevance of studying the paradigm of the interaction of malnutrition (particularly undernutrition) and schistosome-infection, in the face of dynamic and evolving changes in population diets and schistosomiasis epidemiology patterns currently described. This review also intends to validate the use of the outbred and inbred mouse models for investigations on the schistosomiasis/malnutrition association, reporting on recent findings from our laboratory and others about immune response and liver pathology in undernourished mice infected with Schistosoma mansoni. Why study malnutrition in association with schistosome infection? There are several reasons for continuing studies on this association: (i) the frequent overlapping of endemic zones for Schistosoma infection and undernutrition in Brazil and other emerging countries of the developing world, where at least 16% of the total population living in Africa, Asia and Latin America are under the risk of hunger (WFP 2006), (ii) the important role of nutrition in health, in low-income and middle-income countries, estimating that around 32% of the overall disease burden, in terms of mortality and morbidity, would be removed by eliminating malnutrition (Mason et al. 2003), (iii) the concept of parasite-host-environment as interrelated sub-systems, the host nutritional status being included in this context, (iv) the growing importance of the paradigm of the interaction of malnutrition and infection (Solomons 2007), (v) the urbanisation and appearance of new epidemiological profiles of schistosomiasis, emphasising the role of biological, social, economic, political and cultural determinants in the natural history of this helminthiasis, reflecting on the host nutritional status, (vi) the recognition of nutrition as a critical determinant of the immune response, epidemiological and clinical studies stressing that nutritional deficiencies alter immunocompetence (Chandra 1992), which further increase the risk and influence the outcome of infection (Meira 1995, Scrimshaw & SanGiovanni 1997, Fraker et al. 2000, Cunningham-Rundles et al. 2005, Landgraf et al. 2007, Solomons 2007) and, finally, (vii) the need for a deeper evaluation on the real importance of nutrition in schistosome-infected hosts, in view of incomplete and sometimes contradictory results reported in the literature. Schistosomiasis and malnutrition: why is this association still misunderstood? There are several reasons to explain why the interrelationships between schistosomiasis and malnutrition still represent a misunderstood association. In humans: (i) the use of non-standardised and even inadequate methodologies in field investigations, with most papers describing cross-studies or inadequate sample sizes, (ii) the results are randomly chosen, focusing on either the acute or the chronic phase of the disease, (iii) work is done in areas of low prevalence for malnutrition or low intensity of infection and morbidity for schistosomiasis and (iv) generalised conclusions are based only on the clinical form of schistosomiasis locally prevalent, which may influence the type and severity of associated malnutrition. In animal models: (i) the use of diverse experimental animal models, such as mice, rats, gerbils and hamsters, (ii) results are based upon infections with different Schistosoma species and parasite strains, (iii) different methodological approaches are used to induce undernutrition, including mice age at the beginning of the experiment and the use of different experimental diets and (iv) difficulties in applying the results obtained with animal models to humans. Animal models used in experiments on the association schistosomiasis/malnutrition The need for the use of experimental animals and their contribution to elucidate several aspects of human diseases has been recognized a long time ago. Warren (1968) stated: "the elucidation of the pathophysiology and pathogenesis of a disease has rarely been achieved by study of the human patient alone, requiring almost invariably the development of an animal model". Besides being used to reproduce anatomo-pathologic and pathophysiologic features of the infection in humans, laboratory animals have also been frequently utilised for the study of immunity and treatment. In the particular case of studies on the schistosomiasis/malnutrition association, outbred and inbred mice of different lineages have been widely used (Dewitt 1957, Kanuft & Warren 1969, Akpom & Warren 1975, Couti-nho et al. 1991, 1992, 1997, 2003, 2007), but other experimental hosts, such as the albino rat (Bhattacharyya 1965, Parent et al. 1984), the gerbil (Hashem & Fahmy 1962) and the hamster (Mansour et al. 1983) have also been mentioned in investigations on the above association. Liver pathology and schistosomiasis: the importance of the hepatic fibrosis Liver fibrosis is the most important tissue lesion found in schistosome-infected hosts. The chronic granulomatous host response around disseminated parasite eggs, aggravated by fibrosis, is the major contributor to the pathology of the disease (Colley 1981, Smithers & Doenhoff 1982, Warren 1982). T helper (Th) cells (CD4+ T cells) are instrumental for the formation of granulomas around S. mansoni eggs (Mathew & Boros 1986). Moreover, CD8+ T cells appear to be important for down-regulation of granuloma size in chronic infections (Henderson et al. 1992), but antibodies are necessary in addition to down-regulated T cells (Jankovic et al. 1998). Fibrosis enhances the disease pathology, as it is an important feature of advanced schistosomiasis, both in man and in experimental animal models. It represents the anatomical counterpart of the clinical condition known as hepato-splenic schistosomiasis. It follows the granulomatous inflammatory response and occurs mostly at the site of the resolving granulomatous reactions, but fibrosis bands also appear around portal veins distant from granulomas, being partly a sequel to the chronic thrombophlebitis of portal veins. In addition to portal fibrosis, collagen bundles are found in Disse's spaces located underneath the liver sinusoids between the endothelium and the hepatocytes (Boros 1989). Although the pathogenesis of human pipe-stem fibrosis is still not well understood, the host immune response appears to be involved in the overall process of fibrosis (Boros 1989). Thirty to 50% of well-nourished outbred albino Swiss mice subjected to relatively mild (1-2 worm pairs) and prolonged infections (16 weeks or more) with S. mansoni, develop a lesion similar to human Symmer's fibrosis (Fig. 1), as a consequence of a massive and continuous deposition of eggs along the dilated intra-hepatic venous portal branches (Andrade 1987, Andrade & Cheever 1993, Andrade et al. 1997). However, previous study in our laboratory showed that undernourished mice maintained under the same conditions completely failed to develop the "pipe-stem" fibrosis of the liver, unlike the well-nourished controls (Coutinho et al. 1997). Various attempts to induce the pipe-stem like liver fibrosis in undernourished mice have been unsuccessful, such as: shifting from a deficient to a balanced diet and vice-versa (Coutinho et al. 2003); repeated infections five times at 15-day intervals (Coutinho et al. 2007) and assays with BALB/c and C57BL/10 inbred mouse strains (Coutinho 2004). Immune response in well-nourished mice infected with S. mansoni The pathogenesis of schistosomiasis in well-nourished mice is known to be dependent on immunologic phenomena, such as the formation of delayed hypersensitivity-type granuloma around parasite eggs trapped in the liver (Warren et al. 1967). In addition, immunomodulatory mechanisms have been described and appear to be dependent on cell-mediated immunity (Fanning et al. 1981). Most of the host morbidity in schistosomiasis is the result of the parasite antigen-specific host immune responses that are expressed as inflammatory reactions. During the acute phase of the infection, parasite antigens evoke strong humoral and T-cell mediated immune responses, which decrease as the disease advances to chronic phase (Boros 1989), which is conspicuous for well-nourished hosts in experimental murine schistosomiasis. Immune response in undernourished mice infected with S. mansoni Humoral immune response - According to Bastos et al. (1984), the most important immunoglobulins detected in the immune response of well-nourished schistosome- infected mice are IgG1, IgA and IgM, also described in human patients. In Swiss Webster infected mice subjected to a low protein and multideficient diet, Coutinho et al. (1997) and Oliveira et al. (2004) studied the humoral immune response for some antibodies and found that the levels of serum soluble egg antigen (SEA)-specific IgG1, IgG2a, IgG2b and IgG3 were 2-4-fold lower than those found in well-nourished infected controls, probably indicating a generalised low protein nutritional stat-us in these animals. Further experiments using inbred C57BL/6 infected mice led to similar results (Fig. 2) and in addition showed very low levels for IgE in these undernourished animals (Barros 2008). Undernutrition in these animals was induced by the Regional Basic Diet (Coutinho et al. 1997), which produces the clinical, biochemical and pathological features of marasmus in mice, one of the clinical forms of protein-energy malnutrition of humans (Teodósio et al. 1990, Coutinho et al. 1992). Cellular immune response in undernourished infected mice - In the well-nourished host, the immune response in the course of schistosome infection progresses through at least three phases. In the first 3-5 weeks, during which the host is exposed to migrating immature parasites, the dominant response is Th1-like. As the parasites mature, mate and begin to produce eggs at weeks 5-6, the Th1 component decreases and this is associated with the emergence of a strong Th2 response, primarily induced by egg antigens. During the chronic phase of infection, the Th2 response is modulated and granulomas that form around newly deposited eggs are smaller than at earlier times during infection (Pearce & MacDonald 2002). According to Cunningham-Rundles et al. (2005), in the human malnutrition (marasmus) the cellular immune system is more directly affected than the humoral immune system (McMurray et al. 1981, Chandra 1983) and in this situation lymphocyte subsets are altered (Nájera et al. 2004). Malnutrition is also an important cause of immuno-suppression (Peters-Golden et al. 2005). In children, adolescents and young adults infected with Schistosoma japonicum, hepatic fibrosis, the most important lesion found in chronic schistosomiasis, is associated with impaired nutritional status. It was hypothesised that the main mechanism through which hepatic fibrosis leads to undernutrition and anaemia is a systemic increase in the levels of pro-inflammatory cytokines, specifically IL-1 and lL-6 and possibly TNF- α (Coutinho et al. 2005). In the last few years, researchers in different countries have studied the role of nitric oxide (NO) in the immune response of mice infected with S. mansoni. NO, a biological molecular mediator produced in mammalian cells by the enzymatic oxidation of L-arginine mediated by NO synthase, is relevant as an intra and inter-cellular messenger and as a cytotoxin released during various physio-pathologic events, including immunological reactions and inflammation. The inducible NO synthase (iNOS) is one of the three NOS isoforms present in virtually all cells and expressed in response to pro-inflammatory cytokines (IFN- γ , TNF- α , IL-1 β ) and/or microbial products (lipopolysaccharide). In well-nourished mice, NO production increases following the deposition of schistosome eggs in the liver. Overproduction of NO can induce tissue damage and may contribute to morbidity during schistosome infection (Brunet et al. 1999). However, NO appears to have anti-inflammatory or pro-inflammatory properties under different conditions (Kubes 1993). It has also been shown that inhibition of iNOS exacerbates the granulomatous reaction in vitro and this observation lead to the speculation of a probable regulatory role of NO in the development of chronic granulomatous inflammation (Oliveira et al. 1998, Hesse et al. 2001). Despite its potential for tissue damage by direct cytotoxic or cytostatic activity, NO appears to contribute to the maintenance of immunologic homeostasis through T-cell suppression or Th-cell differentiation (Bingisser et al. 1998). Our laboratory is currently conducting studies on the kinetics of IFN- γ , IL-4 and IL-10 in undernourished infected iNOS knockout C57BL/6 inbred mice (Ramos 2007) at different time points (45 and 90 days post-infection). Cytokine assays are performed in culture supernatants of spleen cells after SEA stimulation by the capture ELISA technique. We have found that iNOS-2 knockout mice (KO mice) had diverging immunological profiles when compared to their wild-type controls. In knockout well-nourished infected mice the production of this pro-inflammatory cytokine decreased progressively from the 45th-90th day post-infection, while in knockout undernourished animals we noticed a progressive increase in the level of IFN- γ . Wild-type controls showed a lower secretion of this cytokine and its kinetics was similar and independent of the nutritional status of the rodents (Fig. 3). In both undernourished and well-nourished C57BL/6 KO, histological examination after 45 days of infection showed a mild acute inflammatory exudative reaction around trapped eggs in the liver, compared to the very dense acute inflammatory infiltrate (lymphocytes and polymorphonuclear cells) seen in wild-type controls (Fig. 4A-D). Comparison was being made between Fig 4 A x B and C x D. Although a periovular granuloma can be seen in Fig. 4C (well-nourished iNOS KO), the exudative component is not as remarkable as in Fig. 4D (wild-type well-nourished mouse). Knockout infected mice showed again diverging curves of IL-4 production as infection progressed (around 60 days), undernourished animals displaying ascending titres of IL-4 secretion. The same tendency was found in wild-type animals, but here the higher values were detected in well-nourished mice (Fig. 5). IL-10 detection assays showed a similar kinetics trend, with diverging curves for KO, but for well-nourished animals IL-10 secretion declined as infection matured, while deficient mice remained in an almost linear level of low production (Fig. 6). Experiments on the kinetics of IL-13 have also been performed (Silva 2008) at different time points of the chronic phase in infected undernourished mice (60, 90 and 150 days post-infection). These animals were not iNOS KO. We observed that after a transient peak, IL-13 in undernourished infected mice fell abruptly to very low levels at 150 days post-infection, while well-nourished controls remained in the steady state as the chronic phase became well established (Fig. 7). All of the results are shown as an arithmetic mean of three replicates (3 pools of 3 mice per group), with standard error of mean expressed as error bars. Parasite infections cause malnutrition, impair the host immune response and are affected by host nutrition and immunity. In schistosomiasis, the earliest hepatic granulomas are formed in the Th1 environment with down-regulation of Th1 and up-regulation of Th2 responses six weeks after infection (Todt et al. 2000). Egg-induced granulomas formed during S. mansoni infection are CD4+ dependent cell-mediated hypersensitivity reactions associated with a strong Th2 response and are the principal pathologic manifestations of the disease in mice (Grzych et al. 1991, Lukacs & Boros 1993). Elevated IgE antibody response, although not specific, is a classic clinical sign of the infection by S. mansoni. Nevertheless, there is considerable controversy concerning the role of IgE in the host-parasite relationship based on the work in humans and other experimental models (Hagan 1993, Capron & Capron 1994, Cutts & Wilson 1997, Webster et al. 1997). Controversial findings have also been published concerning the biological role of IgE within the experimental model of the primary murine S. mansoni infection (Amiri et al. 1994). IgE is thought to represent a major defence component against schistosomiasis, presumably by targeting the effector cells against the invading larvae in antibody-dependent cell-mediated reactions. However, primary murine S. mansoni infection is not associated with detectable serum IgE until the worms mature, pair and deposit eggs (Sher et al. 1990). In Brazil, adolescents with high resistance to infection by S. mansoni have specific IgE levels that are 6-8-fold higher than those with low resistance. According to Capron & Capron (1994), in a broader context, IgE response in schistosomiasis and its role in the protective immunity have to be regarded as part of the Th2-dependent regulatory circuit. In malnourished children with parasitic infections, specific IgE response is reduced (Hagel et al. 2003). According to El Ridi et al. (2001), published data taken together indicate that IgE is neither essential nor detrimental to protection against S. mansoni in the primary infection. In human schistosomiasis, the demonstration of IgG deposits in the abnormal Disse's space indicates the involvement of immunological reactions in this pathological alteration (Grimaud et al. 1977). In chronic infections (20 weeks), Th2 responses are blunted (Chensue et al. 1992, Henderson et al. 1992). Borojevic (1992) regards the chronic phase of murine schistosomiasis as predominantly Th1 mediated, largely based on increasing ratio of IgG2a to IgG1 and decreasing eosinophil and IgE levels in the chronically infected mice. IFN- γ , IL-4 and IL-10 exert cross-regulatory effects on the Th1-Th2 balance as IL-4 drives the reaction toward Th2, IFN- γ toward Th1 and IL-10 may inhibit either trend depending on the circumstances (Wynn et al. 1997, 1998, Boros & Whitfield 1998). IFN- γ is a cytokine with important anti-fibrogenic properties (Fox et al. 1991). According to Rumbley et al. (1999), eosinophils form the majority of cytokine producing cells in the granuloma and are the dominant source of IL-4. Anti IL-2 and anti IL-4 have shown to remarkably reduce the size of granulomas forming around injected eggs or antigen-coated beads (Lukacs & Boros 1993, Wynn et al. 1993), but these antibodies have only moderate to minimal effects on the granulomatous inflammation around eggs laid in infected animals (Yamashita & Boros 1992, Cheever et al. 1994). IL-4 and IL-13 largely compensate each other during the formation of hepatic granulomas in infected mice. Thus, only minute granulomas are formed when both are suppressed. Decreased levels of IL-10 have been associated with hepatic fibrosis in human S. mansoni infection (Booth et al. 2004) and are significantly higher in those with severe hepatic fibrosis (grades II and III) in the S. japonicum infected patients (Coutinho et al. 2005). This may reflect a counter-regulatory response to increased levels of pro-inflammatory cytokines. Although a previous study has shown that IL-10 plays a central regulatory role in the pathogenesis of schistosomiasis (Hoffman et al. 2000), its relationship to the nutritional status has not been studied (Coutinho et al. 2005). Hepatic fibrosis in schistosome-infected mice is clearly linked to IL-13 and the Th2 response, although fibrosis sometimes occurs around the formed granulomas in the Th1 milieu (Hesse et al. 2000, Hoffmann et al. 2000). Although there is solid evidence in mice for the role of IL-13 and Type-2 cytokines in the progression of hepatic fibrosis, the contribution of Type-1 and Type-2 associated cytokines to liver fibrosis in humans remains unclear (Abath et al. 2006). These findings and our results on the association schistosomiasis/malnutrition collectively seem to indicate that (i) schistosome-infected undernourished mice are able to develop a humoral immune response, but antibody titres are much lower than in well-nourished controls, (ii) the cellular immune response is not suppressed in undernourished infected mice, but in acute infection IFN- γ and IL-10 cytokines are secreted in smaller amounts than in well-nourished controls, (iii) in iNOS-2 KO, rising levels of IFN- γ , IL-4 and IL-10 cytokines can be detected in nutritionally deficient animals as the infection advances to the chronic phase, but the kinetics of their production seems to be opposite to that seen in well-nourished controls and (iv) the unexpected variation in IL-13 cytokine production observed in infected undernourished mice along different time points of schistosome infection may contribute to delay collagen synthesis in the liver of these animals. This event could explain the inability of deficient mice to develop the periportal liver fibrosis (pipe-stem like murine fibrosis) described in both well-nourished outbred and inbred mice. However, further studies must be pursued to clarify the real meaning of these findings. ACKNOWLEDGEMENTS To Drs Silvia ML Montenegro and Claudia MA Costa, for their valuable participation in different steps of some experiments here reported, to Roni E de Araújo, for helping to prepare the histological sections and morphometry, to Claudia L Cavalcanti, who took care of the animals, and to Simone S de Souza, for secretarial assistance. REFERENCES
Copyright © 2010 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc10063f4.jpg] [oc10063f1.jpg] [oc10063f6.jpg] [oc10063f7.jpg] [oc10063f5.jpg] [oc10063f2.jpg] [oc10063f3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}