 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 105, No. 4, 2010, pp. 380-386 Seric chemokines and chemokine receptors in eosinophils during acute human schistosomiasis mansoni Denise Silveira-LemosI, II, III, IV; Andréa Teixeira-CarvalhoII, +; Olindo Assis Martins-FilhoII; Adriano Luiz Souza-SoaresIII; Pollyanna Castro-SilvaI; Matheus Fernandes Costa-SilvaI, II; Pedro Henrique Gazzinelli GuimarãesI; Helena Barbosa FerrazII; Lúcia Alves Oliveira-FragaV; Mauro Martins TeixeiraIII; Rodrigo Corrêa-OliveiraI ILaboratório
de Imunologia Celular e Molecular + Corresponding author: andreat@cpqrr.fiocruz.br Received 6 January 2009 Financial support: UNDP World Bank/WHO SPRTTD, CNPq (478320/2007-8), FAPEMIG (ATC, OAMF, MMT and RCO are CNPq fellowships) Code Number: oc10067 ABSTRACT The recruitment of circulating eosinophils by chemokines and chemokine receptors plays an important role in the inflammation process in acute human schistosomiasis. Our main focus has been on the plasma chemokines (CXCL8/CCL2/CCL3/CCL24) and chemokine receptors (CCR2/CCR3/CCR5/CXCR1/CXCR2/CXCR3/CXCR4) expressed by circulating eosinophils from acute Schistosoma mansoni infected patients (ACT). Our studies compared ACT patients and healthy individuals as a control group. Our major findings demonstrated a plethora of chemokine secretion with significantly increased secretion of all chemokines analysed in the ACT group. Although no differences were detected for beta-chemokine receptors (CCR2, CCR3 and CCR5) or alpha-chemokine receptors (CXCR3 and CXCR4), a significantly lower frequency of CXCR1+ and CXCR2+ eosinophils in the ACT group was observed. The association between chemokines and their chemokine receptors revealed that acutely infected schistosome patients displaying decreased plasma levels of CCL24 are the same patients who presented enhanced secretion of CCL3, as well as increased expression of both the CCR5 and CXCR3 chemokine receptors. These findings suggest that CCL24 may influence the kinetics of chemokines and their receptors and eosinophils recruitment during human acute schistosomiasis mansoni. Key words: eosinophils - chemokine receptors - chemokines - acute schistosomiasis Chemokines are a group of chemoattractant proteins that induce not only chemotaxis but also activation of target cells (Ulfman et al. 2001). It has been shown that some chemokines influence inflammatory cell infiltration and the cellularity that occurs in the granulomatous response around Schistosoma mansoni eggs. For instance, the chemokines macrophage inflammatory protein-1 α (MIP-1 α /CCL3) and regulated upon activation normal T-cell expressed and secreted (RANTES/CCL5) are produced in a model of granulomatous inflammation in mice and ablation of their activity modulates the size of the lesions (Lukacs et al. 1993, 1994, Chensue et al. 1999, Qiu et al. 2001, Souza et al. 2005). Plasma levels of CCL3 are elevated in chronic schistosomiasis patients and correlate with the level of morbidity (Falcão et al. 2002, Souza et al. 2005); blocking CCL3 activity inhibits the granulomatous response in vitro (Falcão et al. 2002). Booth et al. (2004) recently reported that periportal fibrosis in males with S. mansoni infection is associated with low concentrations of CCL5. Eosinophils represent an important line of defence against parasitic infections (Butterworth 1984). In the parasitic disease caused by exposure to S. mansoni larvae, blood eosinophilia rises due to an increase in interleukin (IL)-5, IL-1, IL-3, granulocyte-macrophage colony-stimulating factor and granulocyte colony-stimulating factor (Clutterbuck et al. 1989, Weller 1992) and this increase is dependent on antigens secreted by eggs through the pores of the shell (Race et al. 1969, Sher et al. 1990). It is well established that an important step in the extravasation of eosinophils is their adhesion to the vascular endothelium, which is mediated by selectins and chemokines (Lampinen et al. 2004). However, the acute eosinophil cellular response against eggs and its contribution to granuloma formation is not well studied (Pacheco & Lenzi 1997). Furthermore, there are no reports in the literature showing alteration in the plasma concentration of chemokines in acutely infected patients. Most of the current knowledge on the role of chemokines in schistosomiasis is derived from mice data. In contrast to experimentation in mice, there is a major difficulty faced by those researchers trying to answer questions in acute schistosomiasis in humans. People living in schistosome endemic areas, where it is easy to find infected patients, are unlikely to develop signs and symptoms related to the acute phase of schistosomiasis (Malhotra et al. 1997, King et al. 1998). In the present study, a group of individuals who have not previously acquired schistosomiasis presented various characterised clinical symptoms of the early phase of S. mansoni infection. This allowed observation of certain immunological alterations in a cohesive group of acutely infected patients. In order to get a better insight into eosinophil participation in acute S. mansoni infection, we analysed the expression of chemokine receptors, namely, CCR2, CCR3, CCR5, CXCR1, CXCR2, CXCR3 and CXCR4 in the blood eosinophils of acutely infected patients and non-infected individuals. We also measured the levels of four different chemokines, CCL2 (MCP-1), CCL3 (MIP-1 α ), CCL24 (eotaxin-2) and CXCL8 (IL-8), which span different cellular specificities, in the plasma of these acute patients. Our findings led us to postulate that CCL24 may influence the kinetics of chemokines and their receptors and leukocyte recruitment during acute human schistosomiasis mansoni.
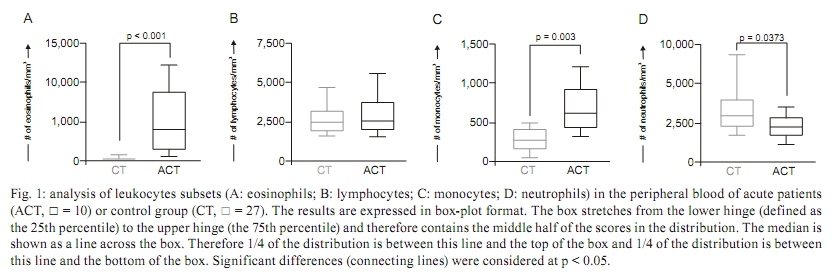
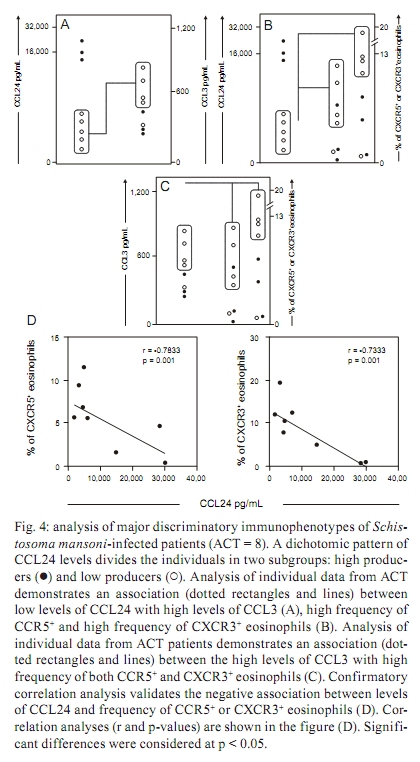
PATIENTS, MATERIALS AND METHODS The study population consisted of two groups of individuals, acute S. mansoni-infected patients (ACT) and healthy blood donors as a control group (CT). The ACT group consisted of 10 patients (6 males and 4 females), with ages ranging from 14-21 years, who acquired their infection in the county village of São Geraldo da Piedade, an endemic area for S. mansoni infection, situated next to the city of Governador Valadares in the state of Minas Gerais (MG), Brazil. In our work, acute infected patients presented various characterized clinical symptoms associated with early infection by S. mansoni, such as fever, diarrhoea, headache, nausea, eosinophilia and, after a few weeks, the presence of schistosome eggs in their faeces. Quantitative parasitological stool examinations and detection of S. mansoni eggs was performed using Kato-Katz (Katz et al. 1972). The ultrasonography analysis did not show echogenicity abnormalities. ACT patients did not receive any corticosteroids or other immunosuppressant chemotherapy, nor were they treated for S. mansoni infection before blood collection. Upon positive laboratory diagnosis of schistosomiasis, all patients received treatment with a single dose of praziquantel, which was independent of their participation in this study. Twenty-seven non-infected individuals (11 males and 16 females) were included as a CT group consisting of volunteer blood donors, aged from 19-30 years and were contacted at the Hemominas Blood Bank Foundation in Belo Horizonte, MG, Brazil. All non-infected volunteers were included after conclusive negative parasitological diagnosis of S. mansoni infection, in addition to negative serology for Chagas disease, leishmaniasis, human immunodeficiency virus infection and hepatitis. The inclusion of all individuals in our investigation was preceded by their written agreement to participate, signed by the patient or by the patient's parents. This study was approved by the Ethical Committees of the Fundação Oswaldo Cruz-Fiocruz, Ministry of Health, Brazil. White blood cell counts - Hemograms were performed in an automated haematology instrument (Coulter MD18, USA), using whole blood collected in 5 mL vacutainer tubes containing ethylenediamine tetraacetic acid (EDTA) as the anticoagulant (Becton Dickinson Biosciences, San Diego, CA, USA). The parameter measured was a differential analysis of leukocyte subsets, including absolute counts of eosinophils, neutrophils, lymphocytes and monocytes. Quantification of plasma chemokines by ELISA - The plasma concentrations of chemokines were measured by sandwich enzyme-linked immunosorbent assay (ELISA), as described by Morita et al. (1999). The ELISA kits for human CCL2 and CCL24 were obtained from R&D Systems, Minneapolis, MN, USA. The antibody pairs and chemokines standards used in the assay for human CCL3 and CXCL8 were purchased from Pharmingen (San Diego, CA, USA). The specific recommendations of the manufacturers were followed. Human plasma samples were subjected to acid-salt precipitation to avoid cross-reactivity in ELISA (Falcão et al. 2002). The concentration of chemokines in plasma was analysed in pg/mL. Immunophenotypic analysis by flow cytometry - Human monoclonal antibodies (mAbs) for anti-CCR2 (48607.211), anti-CCR3 (61828.111), anti-CCR5 (45502.111), anti-CXCR1 (42705.111), anti-CXCR2 (48311.211), anti-CXCR3 (49801.111) and anti-CXCR4 (12G5) were labelled with fluorescein isothiocyanate (FITC) (R&D systems). Anti-mouse IgG1 (679.1Mc7) was used as isotype control and purchased from Becton Dickinson Biosciences Pharmingen (San Diego, CA, USA). Cell staining procedure - One hundred microlitres of EDTA whole blood was incubated in the presence of 5 μ L of undiluted anti-human cell surface molecules mAbs for 30 min in the dark and at room temperature (RT). For CCR2 analysis, monoclonal antibody labelled with biotin (R&D systems, USA) was diluted 1/5 in phosphate-buffered saline (PBS) with avidin-FITC (Diatec, Oslo, Norway) diluted 1/100 in PBS. In this analysis, the cells were incubated with 50 μ L of diluted monoclonal antibody in the dark for 30 min at RT. Then, erythrocyte lysis was performed using 2 mL of FACS Lysing Solution (Becton Dickinson Biosciences Pharmingen, San Diego, CA, USA), followed by incubation for 10 min at room temperature (RT). The leukocyte suspension was further washed with 2 mL of PBS containing 0.01% sodium azide. Prior to flow cytometric acquisition, stained cells were fixed in 200 μ L of FACS fix solution (10 g/L paraformaldehyde, 10.2 g/L sodium cacodylate, 6.65 g/L sodium chloride) for at least 15 min at 4ºC. Data collection from 20,000 events was performed using a FACScan flow cytometer (Becton Dickinson, Mountain View, CA, USA). CELLQUESTTM software provided by the manufacturer was used for data acquisition and analysis. Analysis of eosinophils was performed by single colour immunophenotyping with FL-1/FITC-labelled anti-cell surface marker mAbs. The gating strategy was based on eosinophil selection by their autofluorescent properties in a non-related FL-3 channel vs. forward scatter (FSC) graphs, as described by Weil and Chused (1981). Eosinophil subpopulations were identified using FSC vs. FL3 dot plots. A confirmatory graph was also set up to identify the eosinophils on FSC vs. side scatter dot plot distributions confined to a region of high cell complexity. Data are presented as the percentage of eosinophils expressing a given phenotypic marker, except for CCR3, which was subjected to a semi-quantitative analysis of a single colour histogram of FL1/FITC fluorescence intensity performed as a fluorescence mean channel. Statistics - Differences between groups were first evaluated by MINITAB 13.20 software (San Diego, CA, USA) to examine three parameters, independence, normality and variance, of the data sets. Considering their non-parametric nature, all data sets were analysed using Mann-Whitney's test, with further analysis by Chi-square and Spearman's rank correlation tests using the Graphpad PRISM 4.02 software (La Jolla, CA, USA). Significance was defined in all cases at p < 0.05. RESULTS Acute human schistosomiasis is accompanied by eosinophilia and monocytosis - The assessment of hematological parameters was evaluated in Fig. 1. Data analysis demonstrated that the ACT group displayed a significant increase in the absolute counts of eosinophils as compared to the CT group (Fig. 1A). This eosinophilic profile was accompanied by a significant increase in monocytes (p = 0.003) besides of a decrease in the neutrophils (p = 0.0373) counts in the ACT group as compared to the CT group (Fig. 1C and D, respectively). No significant differences were observed for lymphocyte absolute values (Fig. 1B). Increased plasma levels of alpha/beta-chemokines are found during acute human schistosomiasis mansoni - The ex vivo analysis of the alpha and beta-chemokine patterns in acute patients is illustrated in Fig. 2. Interestingly, data analysis demonstrated a plethora of chemokines secretion with a significant increase in secretion of all chemokines analysed [CCL2 (Fig. 2A), CCL3 (Fig. 2B), CCL24 (Fig. 2C) and CXCL8 (Fig. 2D)] in the ACT group as compared to the CT group. In order to extend our initial findings, we performed a detailed correlation analysis of circulating eosinophil subset counts and chemokine levels. The results of this investigation revealed a negative correlation between the levels of CCL2 and the number of eosinophils (r = -0.64; p < 0.05) (data not shown). No additional significant correlations were found with the additional chemokines investigated. Diminished expression of the alpha-chemokine receptors CXCR1 and CXCR2 is observed during acute human schistosomiasis mansoni - The pattern of alpha and beta-chemokine receptor expression on eosinophils during acute schistosomiasis mansoni is illustrated in Fig. 3. No differences were detected for beta-chemokine receptors, including the percentage of CCR2+ or CCR5+ eosinophils and CCR3 eosinophil expression (Fig. 3A-C, respectively) as well as for the alpha-chemokine receptors CXCR3 and CXCR4 (Fig. 3F, G, respectively). Interestingly, data analysis demonstrated a shift in the alpha-chemokine receptor profile characterised by a significantly lower frequency of CXCR1+ and CXCR2+ eosinophils (p = 0.0458 and p = 0.0084, respectively) in the ACT group as compared to the CT group (Fig. 3D, E, respectively). In addition, a positive correlation between CCR2 and CCR5 expression and eosinophil absolute counts was observed (r = 0.66, p < 0.05; r = 0.74, p < 0.05, respectively) (data not shown). Decreased plasma levels of CCL24 are associated with enhanced secretion of CCL3 and increased expression of CCR5 and CXCR3 in eosinophil subsets during acute S. mansoni infection - The patterns of plasma levels of chemokines and chemokine receptor expression are shown in Fig. 4. Data analysis revealed that ACT patients who displayed decreased plasma levels of CCL24 are the same patients who presented enhanced secretion of CCL3 (Fig. 4A) (p < 0.0001), as well as increased expression of both CCR5 and CXCR3 chemokine receptors (Fig. 4B) (p = 0.002 and p = 0.0337, respectively). In addition, we performed a further investigation that revealed a direct association between CCL3 levels and the percentage of CCR5+ or CXCR3+ eosinophils (Fig. 4C) (p = 0.0001) during acute human schistosomiasis mansoni. In order to confirm the initial findings, we performed a correlation between levels of CCL24 and percentage of CCR5+ or CXCR3+ eosinophils (Fig. 4D). The results demonstrated the existence of a strong negative correlation between levels of CCL24 and the frequency of CCR5 or CXCR3 in eosinophils from acute patients (r = -0.7833, p = 0.001; r = -0.7333, p = 0.001, respectively). Together, these findings led us to postulate that CCL24 may influence the kinetics of chemokines and their receptors, as well as leukocyte recruitment during human acute schistosomiasis mansoni. DISCUSSION Eosinophilia is a hallmark of the acute phase of schistosomiasis - Eosinophils represent a minor cell type among leukocytes, with basal counts ranging from 400-600 cells/mm3 (Brito-Babapulle 2003). In this study, acute schistosomiasis patients had a significantly higher number of eosinophils. Experimental studies have demonstrated that eosinophilia occurs between the 5th-7th weeks after exposure to the parasite and may be induced by Th2 cytokines, such as IL-4, IL-10, IL-13, IL-9 and especially IL-5 (Cara et al. 2000). Hoshi et al. (1995) demonstrated that a transient arrest of rolling eosinophils may be induced by chemokine CXCL8 and shifted into firm long-term arrest by CCL24. Eosinophils then migrate into the tissue in response to chemotactic factors produced locally at the inflammatory site (Teran et al. 1996). Smithers et al. (1977) demonstrated in experimental models that the cellular infiltrate observed in hepatic granulomas is predominantly formed by eosinophils and monocytes. Chemokines in the plasma of acute schistosomiasis patients - The present paper represents the first report of increased chemokine concentrations in the plasma of S. mansoni acute-infected patients. All four of the chemokines measured, CCL2, CCL3, CCL24 and CXCL8, were found at higher levels in the plasma of schistosomiasis patients in the acute phase of the disease as compared to healthy individuals. Although the presence of these chemokines in plasma is not enough to show a definitive role in the manifestations and general immune response associated with acute schistosomiasis in humans, some evidence is available from animal models. CCL24 and CXCL8 are generally known for their chemotactic activity towards eosinophils and neutrophils, respectively. CCL24 is selectively agonistic towards CCR3 receptors, whose expression on eosinophils (Heath et al. 1997), basophils (Uguccioni et al. 1997) and Th2 lymphocytes (Gerber et al. 1997, Sallusto et al. 1998) is compatible with a role in allergic disease and the immune response against helminth infections. In this context, CCL24 could attract eosinophils to inflammatory sites to participate in the initial formation of granulomas. This cell type may comprise up to 40% of the cellular composition of granulomas. Moreover, an increase of CCL24 could indicate a Th2 type immune response. The role of CXCL8 is more evident in acute inflammation, where this chemokine attracts neutrophils to inflammatory sites (Broaddus et al. 1994). Although neutrophils are not obvious contributors to the formation of schistosome granulomatous, this cell type may have a role in the initial influx of cells or at the interface between innate and acquired immunity, collaborating in the secretion of inflammatory mediators that will bring other cells types of greater importance. A decreased frequency of circulating neutrophils in acute schistosomiasis patients may indicate a migration of this cell type into the inflammatory focus. This seems plausible since blood cells from infected patients release mediators in response to soluble eggs antigen stimulation that are chemotactic to neutrophils and eosinophils (Dabes et al. 1989). Moreover, schistosome eggs themselves secrete antigens that have direct chemotactic activity on neutrophils and eosinophils (Owhashi et al. 1985). The chemokines CCL2 and CCL3 are important mediators of the influx of monocytes and macrophages, which are relevant cellular types in the granulomatous response around schistosome eggs. In addition, both macrophages (Koch et al. 1992) and eosinophils (Izumi et al. 1997) are sources of CCL2. Tillie-Leblond et al. (2000) demonstrated that chemotactic activity from eosinophils from bronchial lavage fluid in asthmatic subjects is associated with higher levels of CCL2, CCL3 and CCL5. Moreover, CCL3-deficient schistosome-infected mice exhibit smaller granulomas compared to wild-type mice (Souza et al. 2005). In humans, a correlation between high levels of CCL3 and severity of disease has been observed during schistosome infection (Falcão et al. 2002). Thus, CCL2 and CCL3 could promote the accumulation of macrophages and eosinophils in the early phases of granuloma formation. In our study, both chemokines are increased in plasma from patients with acute S. mansoni infection. Expression profile of chemokines receptors in eosinophils - Signals via chemokine receptors play an important role in the accumulation of eosinophils (Nagase et al. 2001). Previous data have demonstrated a constitutive and as well as or inducible expression of CCR1, CCR3, CXCR1, CXCR2, CXCR3 and CXCR4 by human circulating eosinophils (Heath et al. 1997). In our study, we observed a decrease in the expression of both CXCR1 and CXCR2 by eosinophils from acute patients compared to healthy individuals and a positive correlation between the expression of CCR2 and CCR5 in the ACT group. Schuh et al. (2002), while studying CXCR2 knockout mice during allergic airway reactions and asthma, demonstrated that this chemokine receptor may have significant role in the development and maintenance of both diseases. Heath et al. (1997) demonstrated that after 5-7 days culture in vitro with human IL-5, CXCR2 and to a lesser degree CXCR1 were detectable on the surface of eosinophils and this expression is in parallel with the ability of these eosinophils to migrate to CXCL8 in chemotaxis assays. However, there are conflicting reports on the expression of CXCR2 on circulating eosinophils (Petering et al. 1999, Jinquan et al. 2000, Nagase et al. 2001) and little is known about the expression of chemokine receptors on peripheral blood eosinophils in patients with acute schistosomiasis. Our data suggest that CXCR1 and CXCR2 are not necessary for the activation of circulating eosinophils in the ACT group. Influence of CCL24 on chemokines and their receptors - The study of the importance of chemokines and their receptors for development and maintenance of acute human schistosomiasis has been neglected. Of particular interest are our data describing novel associations between CCL24 and chemokines, such as CCL3, as well as chemokine receptors such as CCR5 and CXCR3. Eotaxin-2 (CCL24) has long been known to be a specific agonist for CCR3, attracting and activating eosinophils, basophils and Th2 type T lymphocytes and its expression profile coincides with a potential role in allergic inflammation or parasitic diseases as schistosomiasis (Petkovic et al. 2004, Silveira-Lemos et al. 2008). Recently, it was reported that eotaxin-3 (CCL26) acts as a natural antagonist on CCR1, CCR2 and CCR5 receptors (Ogilvie et al. 2003, Petkovic et al. 2004) and may play an unrecognised role in the polarisation of cellular recruitment by attracting Th2 lymphocytes, eosinophils and basophils via CCR3, while concomitantly blocking the recruitment of Th1 lymphocytes and monocytes via CCR1, CCR2 and CCR5. Our data showed a strong negative correlation between levels of CCL24 and expression of CCR5 and CXCR3, suggesting that a similar mechanism may be occurring during acute schistosomiasis. In this context, the increased responsiveness of eosinophils from acute patients to CCL3 may be mediated by their specific receptors, such as CCR5. Oliveira et al. (2002) investigated the regulation of chemokine-mediated responses and receptor expression on eosinophils from mice and reported that CCL3 and eotaxin-1 (CCL11) induced a significant intracellular calcium flux in antigen-elicited and peripheral blood eosinophils and that CCL2, CCL22, CCL4 and CCL1 did not. Peripheral blood eosinophils migrated toward CCL3 and CCL11 but did not migrate toward CCL2, CCL22, CCL4 or CCL1. These results suggest that eosinophils may up-regulate and use additional chemokine receptors during the progression of inflammatory and stimulus responses for migration and activation and that the chemokine milieu is very important in this process. Overall, our data demonstrated that the acute phase of schistosomiasis is characterised by an eosinophilic profile that shows the complex relationship between chemokines and their receptors and that it may affect the distribution and recruitment of immune cells towards an inflammatory focus as well as determine the establishment of distinct patterns of immunological parameters during human acute schistosomiasis mansoni. ACKNOWLEDGEMENTS To the technical staff of the Laboratório de Imunologia Celular e Molecular, Fundação Oswaldo Cruz-Fiocruz, and the Núcleo de Pesquisa em Imunologia, UNIVALE, Brazil, for invaluable assistance during this study, and to the PDTIS-Fiocruz, for use of its facilities. REFERENCES
Copyright © 2010 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc10067f3.jpg] [oc10067f2.jpg] [oc10067f4.jpg] [oc10067f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}