 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
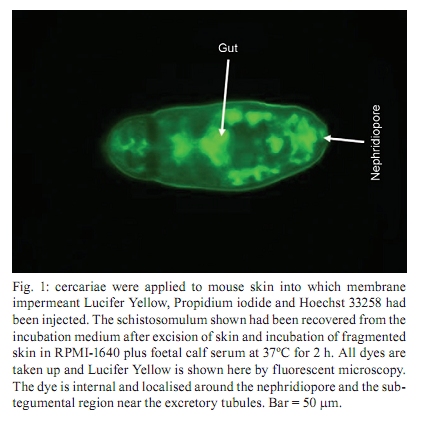
Memórias do Instituto Oswaldo Cruz, Vol. 105, No. 4, 2010, pp. 387-390 Uptake of macromolecules by cercariae during skin penetration and transformation to schistosomula (Schistosoma mansoni) Joyce ThornhillI; John KuselI, +; Fabricia Alvisi de OlivieraII; Fabio RibeiroII; Symone Fulgêncio LimaII; Paulo Marcos Zech CoelhoII; Paul McVeighIII; Ana Carolina Alves MattosII IUniversity
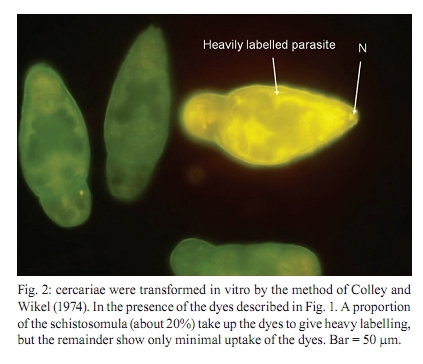
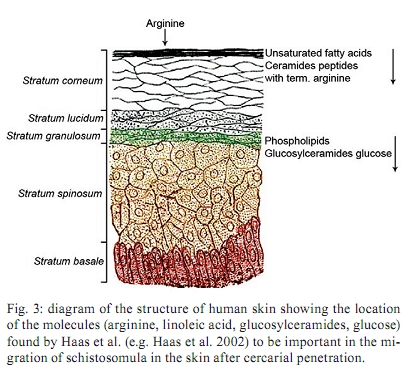
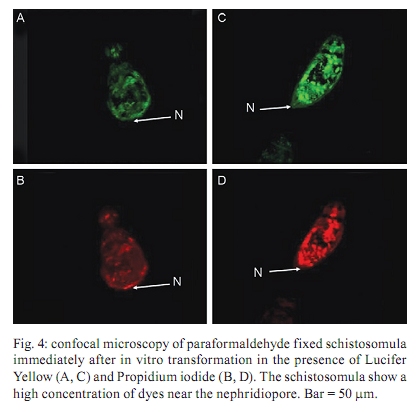
of Glasgow, GBRC 120 University Place, G12 8TA Glasgow, Scotland, UK + Corresponding author: johnkusel@hotmail.com Received 12 November 2008 Code Number: oc10068 ABSTRACT Here, we observed the uptake of membrane-impermeant molecules by cercariae as they penetrate the skin and are transformed into schistosomula. We propose that membrane-impermeant molecules, Lucifer Yellow, Propidium iodide and Hoechst 33258 enter the parasite through both thenephridiopore and the surface membrane and then diffuse throughout the body of the parasite. We present a hypothesis that the internal cells of the body of the schistosomulum represent a new host-parasite interface, at which skin-derived growth factors may stimulate receptors on internal membranes during transformation of the cercariae into the schistosomulum. Key words: Schistosoma mansoni - membrane impermeant Lucifer Yellow - nephridiopore - host/parasite interface - linoleic acid The penetration of mammalian skin by cercariae of Schistosoma mansoni is accompanied by many changes in the structure and metabolism of the parasite (McLaren & Hockley 1976, Skelly et al. 1993, Bahia et al. 2007). A very dramatic change is the loss of the tail via its severance from the body to yield the schistosomulum (Stirewalt et al. 1966, Howells et al. 1975, McLaren & Hockley 1976). During the tail loss, the nephridiopore (the aperture representing the external exit of the excretory system) is fully exposed to the external environment. Recently, a pathway was described that involves the entry of large molecules into the body through the nephridiopore after surface membrane damage (Tan et al. 2003). In the present paper we describe the process of uptake of membrane-impermeable fluorescent dyes by the cercariae as they penetrate the skin and we propose that these molecules may enter through both the nephridiopore and the surface membrane. MATERIALS AND METHODS Fluorescent chemicals - Lucifer Yellow CH lithium and potassium salt, Hoechst 33258, Propidium iodide and Texas Red bovine serum albumin were purchased from Invitrogen (UK). FITC-labelled dextrans with molecular weights of 4, 10 and 70 kDa were purchased from Sigma (UK). All fluorescent chemicals are water soluble and were dissolved (1 mg per mL) in RPMI-1640. Origin of parasite material, skin penetration and in vitro culture - A Puerto Rican isolate of S. mansoni was maintained in our laboratory in Biomphalaria glabrata and the outbred TO strain of mice (obtained from Glasgow University). Skin schistosomula were obtained by allowing cercariae, which were concentrated by light, to penetrate into a shaved area of a recently euthanised mouse. Mice were euthanised by cervical dislocation and their abdomens were shaved with an electric shaver. Two hundred-microlitre intradermal injections containing 1 mg per mL of fluorescent dye were delivered (26 G needle) and flanged open tubes were placed on the skin. The flanges were pressed onto the skin surface. One millilitre of cercarial suspension was gently poured through the open end of tube and, in this way, the cercariae could contact the skin in the region labelled with the dye. After 15 min, the skin region exposed to the cercariae was removed with scissors, cut into small pieces and incubated for 2 h in RPMI-1640 supplemented with 10% foetal calf serum at 37ºC. Production of schistosomula in vitro - Schistosomula were produced in vitro according to the method of Colley and Wikel (1974). Cercariae were suspended in RPMI-1640 during the syringe transformation. Fixation of cercariae - Labelled schistosomula were fixed for 4 h with 4% paraformaldehyde. The 4% paraformaldehyde was prepared by dissolving the solid paraformaldehyde (Sigma-Aldrich, UK) in RPMI-1640 at 60ºC. Examination of parasites by fluorescence microscopy - Photographs were taken after observation of the parasites under a 40X objective lens of a Leica Laborlux S microscope. All labelled parasites were viewed and photographed using a Leitz Orthoplan Laborlux S microscope with a Leica camera attachment. This allowed the variability in the labelling of the parasite population to be observed, counted and recorded. Photographs were taken after observation of the parasites under the 40X or 50X objectives. Further detail in the distribution of the labelled compounds was obtained using a Zeiss microscope (Axioplan 2). Confocal microscopy was carried out with a Leica TCS NT AOBS SP2 confocal microscope. RESULTS In vivo labelling during penetration of fluorescently-infiltrated mouse skin - Schistosomula were recovered from the skin labelled with fluorescent dyes (Lucifer Yellow and Propidium iodide) after an incubation of 30 min or 2 h. The worms were then examined by fluorescence microscopy (Fig. 1). Fig. 1 shows that Lucifer Yellow (0.45 kDa) clearly labelled the subtegumental area close to the excretory tubules, as well as some other organs (e.g., the gut and the nephridiopore). Fig. 1 summarises the results at both 30 min and 2 h. A similar pattern is observed when fluorescent dextrans with molecular weights of 3-20 kDa were used in the skin assay (results not shown). Transformation of cercariae into schistosomula by syringe passage - Cercariae were syringe transformed into schistosomula in the presence of each of the dyes. A subsequent incubation at 37ºC in RPMI-1640 showed that, although some of the schistosomula incubated in Lucifer Yellow appeared very similar to the skin transformed schistosomula (Fig. 1), the results were much more variable for these mechanically transformed forms and only a proportion of the parasites had taken up the dye to provide a clear internal localisation. This result is illustrated in Fig. 2, which shows that many of the schistosomula were diffusely fluorescent, without the clear definition of organelles and tissues apparent in Fig. 1. In Figs 1 and 2, free swimming cercariae were incubated at RT with Lucifer Yellow for 30 min and washed three times in RPMI-1640 as a negative control. Some Lucifer Yellow adhered to the surface and very small amounts entered the body, providing a very diffuse pattern unlike the definite structures observed in Fig. 1. In a few specimens, the fine outline of the excretory tubules could be seen. Any internal Lucifer Yellow was excreted from these parasites (data not shown). Specific stimulus from skin - The variability in the amount of fluorescent label (Lucifer Yellow and Propidium iodide) taken up by the cercariae appeared to be much less in those parasites recovered from the skin after 30 min or 2 h, whereas in vitro-transformed schistosomula showed considerable differences in uptake between individual parasites. These differences were not genetically determined since three clones of cercariae, produced from monomiracidial infections of snails, also displayed this variability. When the brightly labelled parasites (such as that seen in Fig. 2) were counted and expressed as a percentage of the total population after transformation in the presence of the dyes, the values obtained for the three clones were 11%, 14% and 24%. The value obtained for a mixed population of cercariae (from a routine multi-miracidial infection) was 17%. We argue here that there must be a specific stimulus from the skin penetration process that was lacking in the in vitro transformation process. The composition of the medium used during syringe transformation was varied in an attempt to reduce the differences in uptake between parasites formed in vitro and those formed on the skin. Uptake occurred to the same extent (between 10-25% brightly labelled parasites) in RPMI-1640, sodium chloride (0.15 M), potassium chloride (0.15 M) and choline chloride (0.15 M), but it was inhibited (5% brightly labelled parasites) in glucose (0.2 M in water) and in water alone. From these results, we concluded that some kind of charged molecule must be present in the transformation fluid to allow uptake to occur. When the cercariae were transformed in 50% foetal calf serum in RPMI-1640, the percentage of parasites that were brightly labelled increased to 60%. Thus, foetal calf serum may contain a compound that stimulates uptake. We conclude that there is a specific, but as yet unknown, stimulus provided to the cercariae from the skin that enables the cercariae to open the nephridiopore or alter the surface membrane permeability and allow the uptake of molecules. Nature of the skin stimulus - The work of Haas et al. (1997, 2002) and Grabe and Haas (2004) provided insight as to the nature of the skin stimulus. At the epidermal surface, arginine, ceramides and linoleic acid stimulate the parasite, while phospholipids, glucosyl ceramides and glucose are the major stimuli deeper in the epidermis (Fig. 3). Two hours after the transformation, linoleic acid (0.5 mM) was added to the schistosomula for a period of 15 min in the presence of Lucifer Yellow. The results showed a highly significant increase in the number of brightly fluorescent schistosomula which showed uptake of the fluorescent dyes (in a mean of 6 experiments), with an increase from 16% brightly labelled parasites (without linoleic acid ) to 75% brightly labelled parasites in the presence of linoleic acid. Whether this uptake occurs through the nephridiopore or through the surface membrane has yet to be determined. Evidence that uptake of fluorescent dyes might occur through the nephridiopore - Cercariae transformed in vitro in the presence of Lucifer Yellow were fixed in paraformaldehyde and examined by confocal microscopy. In the brightly labelled parasites, the major area of dye localisation was near or in the nephridiopore (Fig. 4). However, in many images, the surface membrane was also labelled. Thus, uptake may also occur through the surface membrane. Cercariae were incubated with Propidium iodide and Hoechst 33258 at RT in vitro and watched under a fluorescence microscope. Over a period of 20 min, the dyes were initially observed at the nephridiopore and they then could be seen to diffuse into the body of the cercaria and follow a pathway adjacent to the excretory tubules (Fig. 5). Since these experiments were carried out at RT, we cannot exclude the possibility that the region of the surface membrane near the nephridiopore is permeable to these dyes. DISCUSSION In this paper, we report that, during skin penetration or syringe transformation of cercariae, membrane-impermeant molecules such as Propidium iodide and Lucifer Yellow can enter the body of the parasite. FITC dextrans (3-20 kDa) were also observed to enter the body of the parasite in a similar fashion to Lucifer Yellow (data not shown). These molecules may enter the parasite through the nephridiopore or the surface membrane. This entry of membrane impermeant molecules may have considerable implications. If skin-derived amino acids (Meyer et al. 1991) enter the parasite, they may have a nutritive function. Skin-derived growth factors have a molecular weight (6-22 kDa) that might enable them to enter the parasite. Epidermal growth factor (Shirakata et al. 2003), keratinocyte growth factor (Escámez et al. 2004) and insulin-like growth factor-1 (Jeschke & Herndon 2007, Todorovic et al. 2008), following entry into the parasite, may activate receptors in the parasite and affect its growth and development. Receptors for all these growth factors are present in the schistosome genome (www.schistoDB.net). We envision a scenario in which growth factors from the skin enter the tissues of the schistosomulum and stimulate changes in tegument metabolism and synthesis. This hypothesis can be tested by comparing the transcriptome and proteome of in vitro-generated and skin-derived schistosomula and examine the changes in signalling pathways (Dissous et al. 2007, Loverde et al. 2007, Walker & Rollinson 2008). ACKNOWLEDGEMENTS John Kusel thanks the Leverhulme Trust for a travelling grant to attend the XI Symposium of Schistosomiasis honouring Dr. Piraja da Silva. REFERENCES
Copyright © 2010 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc10068f2.jpg] [oc10068f5.jpg] [oc10068f3.jpg] [oc10068f4.jpg] [oc10068f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}