 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
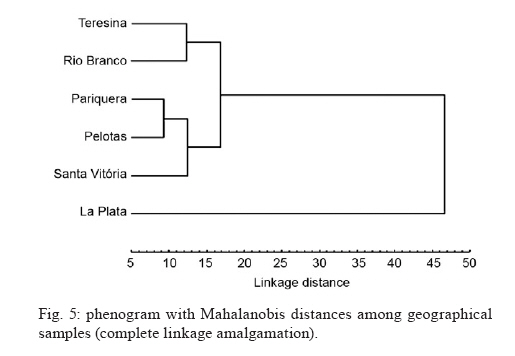
Memórias do Instituto Oswaldo Cruz, Vol. 105, No. 5, 2010, pp. 672-676 ARTICLES Genetic-morphometric variation in Culex quinquefasciatus from Brazil and La Plata, Argentina Sirlei Antunes de MoraisI; Camila MoratoreII, III; Lincoln SuesdekII, III; Mauro Toledo MarrelliI, + IDepartamento
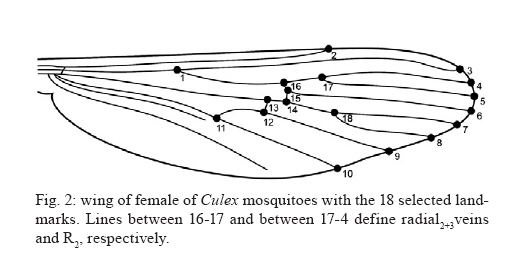
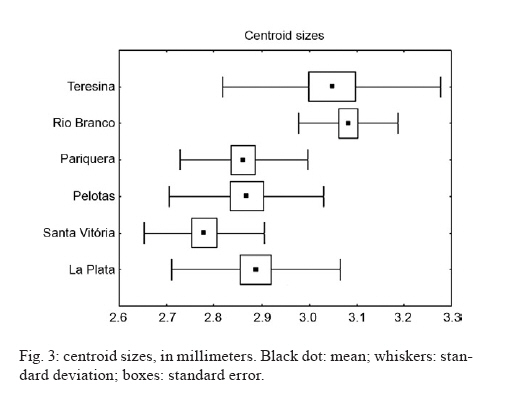
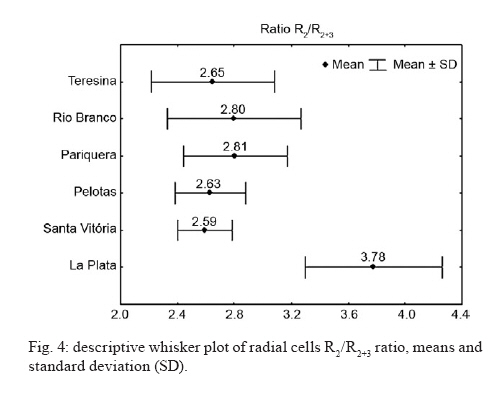
de Epidemiologia, Faculdade de Saúde Pública + Corresponding author: mmarelli@usp.br Received 24 November 2009 Code Number: oc10113 ABSTRACT Variation among natural populations of Culex (Culex) quinquefasciatus Say is associated with different vectorial capacities. The species Cx. quinquefasciatus is present in the equatorial, tropical and subtropical zones in the Brazilian territory, with intermediate forms between Cx. quinquefasciatus and Culex pipiens occurring in regions of latitudes around 33°-35°S. Herein, we studied geographically distinct populations of Cx. quinquefasciatus by genetic characterization and analysis of intra-specific wing morphometrics. After morphological analysis, molecular characterization of Cx. quinquefasciatus and intermediate forms was performed by polymerase chain reaction of the polymorphic nuclear region of the second intron of the acetylcholinesterase locus. Additionally, the morphology of adult female wings collected from six locations was analyzed. Wing centroid sizes were significantly different between some geographical pairs. Mean values of R2/R2+3 differed significantly after pairwise comparisons. The overall wing shape represented by morphometric characters could be divided into two main groupings. Our data suggest that Brazilian samples are morphologically and genetically distinct from the Argentinean samples and also indicated a morphological distinction between northern and southern populations of Brazilian Cx. quinquefasciatus. We suggest that wing morphology may be used for preliminary assessment of population structure of Cx. quinquefasciatusin Brazil. Key words: Culex pipiens complex - Culex quinquefasciatus - clines - molecular identification - wing morphology The Culex (Culex) pipiens complex in the Americas is composed of two main species, Culex (Culex) quinquefasciatus Say, which is adapted to tropical zones and Cx. pipiens L., which is found in temperate zones. In intermediate areas, these two members can mate yielding hybrids and intermediate forms (Barr 1957), through the processes of gene flow and introgression (Humeres et al. 1998, McAbee et al. 2008, Kothera et al. 2009). Mosquitoes of the Cx. pipiens complex are potential vectors of filarid worms in tropical and subtropical areas (Bockarie et al. 2009) and diverse arboviruses, including West Nile virus (Apperson et al. 2004, Cook et al. 2006). Adult females and immature stages of the Cx. pipiens complex are morphologically similar. The differentiation of adult males can be made by analysis of genitalia, which includes measuring the DV/D ratio of the dorsal arms of the aedeagus in the phallosome, as reported by Sundararaman (1949). Because members of the Cx. pipiens complex have a broad geographical distribution, other phenotypical differences beyond those of the male genitalia can be found. For example, some wing characters can also been used for morphologic identification of mosquito species. The taxonomic keys compiled by Forattini (2002) compared wing morphology of adult females. In that report, the subcosta vein (Sc) was reported to bind with the costa vein (C) before the bifurcation of the radial2+3 vein (R2+3) in Cx. quinquefasciatus, whereas in Cx. pipiens, the Sc vein binds to vein C at the same point or beyond the R2+3 bifurcation. To our knowledge, these findings have not been investigated in other geographically distant populations. Linam and Nielsen (1962) reported that Bekku (1956), in Northern Japan, also attempted to use wing characteristics [the length of radial cell R2 (second marginal cell) divided by the length of vein R3] to distinguish adults of the pipiens group. The results of Bekku (1956) were inconclusive, which was later explained when Kamura (1958) found that these particular wing measurements varied with the temperature of the environment. Nielsen and Rees (1961) and Linam and Nielsen (1962) reported that the most reliable method of separating females of Cx. pipiens and Cx. quinquefasciatus was by a wing measurement value obtained by dividing the length of the cell R2 by the length of the vein R2+3 (R2/R2+3 ratio). These authors reported the value of this ratio to be about 5.0 in Cx. pipiens and 3.0 or less in Cx. quinquefasciatus, with intermediate values present in the hybrids. However, these studies did not evaluate morphometric data among mosquito populations and climatic/geographic gradients. In insects, the phenotypic variations among members of species complexes have been intensely investigated, especially those concerning the size and shape of adult wings (James & Partridge 1998, Morin et al. 1999). In Drosophila, which has a worldwide distribution, significant morphometric wing variations were found in populations from different geographic regions, forming clusters or latitudinal clines (Van'T Land et al. 1999). Some studies report that variations in the wings may be related to intrapopulation genetic variations, life history [such as conditions during larval development (Swindell & Bouzat 2006)], the direction of selection throughout the population sources (Hansen & Houle 2008) and the fitness or possible targets of natural selection (Buggs 2007, Gay et al. 2008). Other studies have related morphological differences to species divergence or ecological adaptations (James et al. 1997). The taxonomy of the Cx. pipiens complex is difficult to interpret, since members of the complex present several similarities in terms of both genetics and morphology. To correctly distinguish the taxa, morphological and molecular investigations are needed, as well as knowledge about the characteristics of populations throughout their biogeographical gradients. Likewise, the correct identification of females and hybrids is of epidemiological importance, mainly due to ecophysiological differences, which include autogeny, reproductive diapause, stenogamy, feeding behavior and vector potential (Humeres et al. 1998, Lenormand & Raymond 2000, Fonseca et al. 2004, Savage et al. 2008). Molecular markers have become an important tool to solve taxonomic problems and to identify sibling species. For example, Bourguet et al. (1998) reported variability at the nucleotide level of the acetylcholinesterase (ace-2) gene in several strains and natural populations of the Cx. pipiens complex. Few polymorphisms were found in coding regions within a subspecies, but many polymorphisms were observed between subspecies in noncoding regions. In their study, Bourguet et al. (1998) described a method based on a restriction enzyme polymorphism in polymerase chain reaction (PCR) amplified DNA, in which the presence or absence of a single restriction site discriminates between Cx. pipiens, Cx. quinquefasciatus and their hybrids. This technique has since been used to reliably discriminate between mosquitoes from more than 30 worldwide strains or populations. The present paper is a preliminary investigation of genetic-morphological variability of Cx. quinquefasciatus in Brazil. In addition, the presence of a hybrid zone for Cx. quinquefasciatus and Cx. pipiens in Argentina is discussed. It also revises the morphological identification data of the females by morphometric characters of the wing. MATERIALS AND METHODS Mosquito samples - Immature stages of mosquitoes were collected in the field. Larvae were reared to adults in the laboratory and examined after emergence. The samples were collected in urban areas during the hot months of February and March of 2008, in the Brazilian municipalities of Teresina (5°05'S), Rio Branco (9°58'S), Pariquera-Açu (24°42'S), Pelotas (31°45'S) and Santa Vitória do Palmar (33°31'S). Samples were also collected in the city of La Plata (34°55'S), Argentina (Fig. 1). Sampling covered a latitudinal extension of 30° throughout South America. Mosquitoes were identified as Cx. quinquefasciatus by analysis of morphologic characters, according to taxonomic keys in Forattini (2002). For wing morphology and molecular analyses, only females were selected because species diagnosis is more difficult among female adults. Each individual was used for both analyses. DNA was extracted from whole bodies (without wings). Additionally, adult males were used for description of genitalia (phallosome) and calculation of DV/D ratio (Sundararaman 1949). Nucleic acid isolation - After removing of the wings for morphometric analysis, genomic DNA was isolated from the remaining body parts using Qiagen DNeasy® Blood & Tissue kits, following the manufacturer's recommendations with slight modification. DNA was eluted in a final volume of 100 μL and stored at -20°C until used for PCR. PCR - Taxon-specific PCR primers based on polymorphisms in the second intron of the ace-2 locus were used to assist in distinguishing sibling species of Cx. pipiens complex and to characterize populations and zones of hybridization (Bourguet et al. 1998, Aspen & Savage 2003, Smith & Fonseca 2004, Savage et al. 2007, Sanogo et al. 2008, Kothera et al. 2009). The fragment of the ace-2 locus was amplified from each DNA sample by PCR in a final reaction volume of 50 μL with 2 mM Mg++, 20 mM Tris-HCl (pH 8.4), 50 mM KCl, 0.5 mM of each primer, 0.2 mM dNTP mix, 2 U of Taq DNA polymerase (Invitrogen) and approximately 6.0 ng of genomic DNA. We used multiplex forward and reverse primers for each species of interest: ACEquin 5'-CCTTCTTGAATGGCTGTGGCA-3', ACEpip 5'-GGAAACAACGACGTATGTACT-3', B1246s 5'-TGGAGCCTCCTCTTCACGG-3'. All primers were described by Smith and Fonseca (2004). The multiplex produces a 610-bp fragment in Cx. pipiens, a 274-bp fragment in Cx. quinquefasciatus and may produce one or both fragments in hybrids of the two species. The amplification program consisted of one cycle at 94°C for 5 min, followed by 35 cycles at 94°C for 30 sec, 55°C for 30 seconds, 72°C for 1 min and one cycle at 72°C for 5 min. Wing morphology - Wings of females from the following six locations were analyzed: Teresina (n = 22), Rio Branco (n = 27), Pariquera-Açu (n = 28); Pelotas (n = 22); Santa Vitória (n = 22) and La Plata (n = 31). Right wings were mounted on slides under a coverslip using Entellan. Photographs of wings were taken with a Leica DFC320 digital camera coupled to a stereomicroscope Leica S6 using 40X magnification. In each wing image, coordinates of 18 landmarks (Fig. 2) were digitized using tpsDig software (v1.40 James Rohlf). Subsequent morphological analyses were performed according to Rohlf (1993). Computation of the R2/R2+3 ratio for each individual was performed using landmarks 4, 16 and 17, according to Nielsen and Rees (1961) and Linam and Nielsen (1962). Scores of this ratio were statistically compared within and among the six geographical samples. Overall wing shape based on the 18 landmarks was compared among geographical samples through discriminant multivariate analysis. Following this analysis, we generated validated reclassification tests and a phenogram of Mahalanobis distances within each pair of samples. For overall wing size assessment, we used the centroid size as extracted from coordinates of the 18 landmarks. Morphometric analyses were performed using tpsRelw 1.44 (Rohlf 2006) and PAD softwares (Dujardin 2002) and graphical outputs were generated using Statistica 7.0 software (StatSoft). RESULTS Our results confirmed that Cx. quinquefasciatus is distributed in the equatorial, tropical and subtropical zones in the Brazilian territory, with hybrids forms between Cx. quinquefasciatus and Cx. pipiens occurring in regions of latitudes around 34-35°S, in La Plata. The multiplex PCR produced a 274-bp fragment in all specimens from Brazilian localities (Teresina, Rio Branco, Pariquera, Pelotas and Santa Vitória), identifying them as pure Cx. quinquefasciatus populations. In the sample from La Plata (Argentina), three PCR band profile were found: (i) only the 274-bp band (35% of specimens), (ii) only the 610-bp band (23%) and (iii) both bands (42%). These results suggest that La Plata is a hybrid region. Therefore, in this study, the samples collected in the Brazilian regions were labelled Cx. quinquefasciatus (Q) and the samples from La Plata were labelled as a population of hybrid origin (H). In another study (unpublished observations), 3% of the samples collected from Santa Vitória do Palmar in the winter showed fragments with both bands of ace-2. This suggested that the border of an area with high levels of hybridization area is approaching the latitude of 33°S, in Brazil. Our findings are in accordance with the observations by Almirón et al. (1995) and Humeres et al. (1998), which indicated the presence of hybrids in the region of Argentina based on morphology of male genitalia, allele frequencies and experimental crosses. Those authors suggest latitudes between 33-35°S as a possible hybridization region. Among Brazilian samples, populations from the tropical locations Teresina and Rio Branco had larger wings than subtropical areas (Pariquera, Pelotas and Santa Vitória do Palmar). Pairwise comparisons of samples from the six geographical regions using non-parametrical Kruskal-Wallis + Dunn's post-hoc test indicated significant differences in centroid size (p < 0.05) between Teresina and the others (except Rio Branco) and between Rio Branco and the others (except Teresina) (Fig. 3). No other pairs of samples differed significantly regarding the centroid size. These results suggest a temperature-depended cline. Other studies have suggested that unlike wing shape, wing size may be a labile trait influenced by temperature (James et al. 1997, Gibert et al. 2004, Polak et al. 2004). However, it is currently difficult to perform a more profound interpretation of wing size data because wing size in culicids is reportedly also influenced by food availability (Jirakanjanakit et al. 2007). The R2/R2+3 ratio scores of Brazilian samples were in accordance with the Cx. quinquefasciatus geographical zones, whereas the scores of Argentinean samples were similar to mosquitoes of hybrid origin, reported by Linam and Nielsen (1962) (Fig. 4). Comparisons using non-parametrical Kruskal-Wallis + Dunn's post-hoc test indicated that samples from La Plata differed significantly (p < 0.001) from samples of the other five geographical regions. No other pair of samples differed significantly regarding that ratio. These results may be indicative of low gene flow between Q and H populations because wing shape is genetically determined and has evolutionary significance (Dujardin 2008, Jirakanjanakit et al. 2008). A possible barrier to gene flow between Q and H would be the geographic distribution of Cx. pipiens, which appears to be climatically constrained to south of 33ºS, according to Almirón et al. (1995) and Humeres et al. (1998). Discriminant analysis of wing shape data obtained from the 18 landmarks yielded a matrix of Mahalanobis distances, which is represented as a phenogram in Fig. 5. The overall wing shape showed a clear distinction among the Brazilian (Q) and Argentinean (H) samples, with two visible major clusters. Validated reclassification tests also showed that these two major clusters can be distinguished with 100% accuracy. Among Brazilian samples, there is an inner division separating northern (Teresina + Rio Branco) from southern (Pariquera + Pelotas + Santa Vitória) geographical samples. Genetic data based on ace-2 amplification corroborated the morphological divergence between populations of Brazilian and Argentinean mosquitoes. Populations from La Plata did not show a homogeneous banding pattern, indicating that this population may be composed of hybrids that originated from an introgression event between Cx. quinquefasciatus and Cx. pipiens species. DISCUSSION Our data suggest that the Brazilian populations of Cx. quinquefasciatus sampled are morphologically and genetically distinct from the Argentinean population of La Plata. These results, which are similar to some observed in North America (Linam & Nielsen 1962), indicate that wing characters may serve as diagnostic markers in future studies of the pipiens complex. However, more studies will certainly be needed to confirm such usefulness and to determine the geographical extension of the hybrid zone around La Plata. Wing shape analysis also revealed a geographical population structure in the Brazilian samples. Northern samples, collected from a tropical zone, clustered separately from southern populations (collected from a subtropical zone), which leads us to believe that gene flow is lower between these two regions than within them. This conclusion is supported by the knowledge that wing shape can be indicative of population genetic structure since it is strongly determined by quantitative genetic heritage (Dujardin 2008). Morphological divergence between northern and southern samples might be enhanced by a geographical barrier, since the shortest linear distance between any pair of northern-southern locations (Teresina - Pariquera) is over 2100 km. A morphoclimatic barrier consisting of the transition between tropical and subtropical zones is another hypothesis to be tested. Genetic data were homogeneous for the five Brazilian samples regarding this portion of ace-2, suggesting that Cx. pipiens is not present in those Brazilian locations. Male representatives of the three main wing morphological clusters exhibited the following DV/D mean ratios: Q-northern (Rio Branco) = 1.02; Q-southern (Pelotas) = 0.86 and H (La Plata) = 0.02. Although these results are preliminary (sample sizes were 2, 4 and 2, respectively) and part of another survey in course, they are in accordance with the expected DV/D ratios: values near to zero are typical of Cx. pipiens (Sundararaman 1949). In addition, the overall shape of phallosomes of H samples and H samples were more similar to Cx. quinquefasciatus and Cx. pipiens, respectively (data not shown). Thus, data from males reinforces the evidence that population H represents a hybrid zone. Differences between northern and southern Brazilian samples suggest that wing characters are also useful for a preliminary assessment of population structure of Cx. quinquefasciatus in Brazil and should be helpful if utilized in population genetic surveys. ACKNOWLEDGEMENTS To Dr. Almério de Castro Gomes (School of Public Health, University of São Paulo), for providing mosquito samples from Teresina, to Gustavo Rossi (Centro de Estudios Parasitológicos Y de Vectores, La Plata, Argentina), for providing specimens from La Plata, and to Ministry of Health (FUNASA) and Departments of Health of municipalities of Pelotas and Rio Branco, for helping us during the field works. REFERENCES
Copyright © 2010 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc10113f1.jpg] [oc10113f3.jpg] [oc10113f5.jpg] [oc10113f4.jpg] [oc10113f2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}