 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
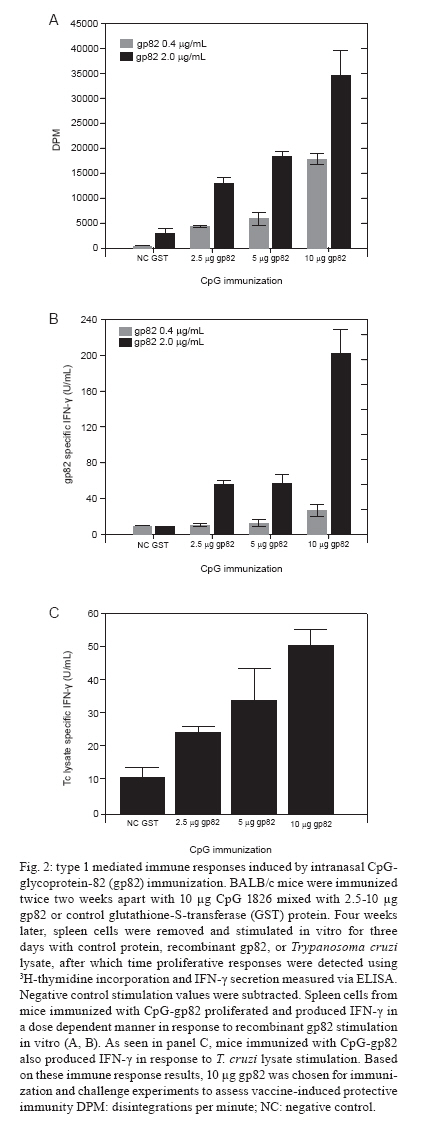
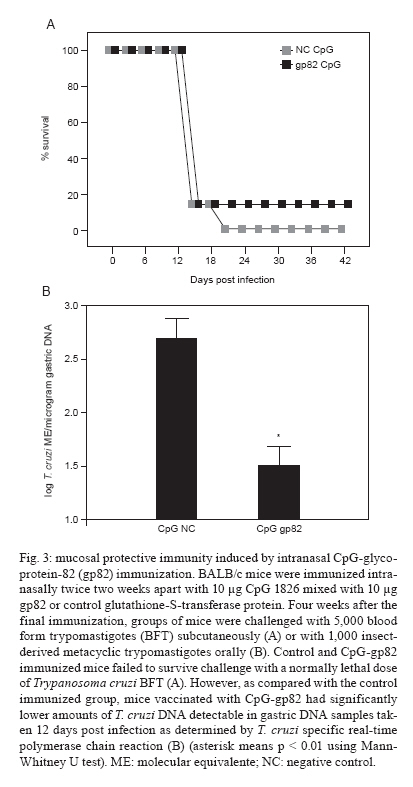
Memórias do Instituto Oswaldo Cruz, Vol. 105, No. 5, 2010, pp. 687-691 ARTICLES Immune responses to gp82 provide protection against mucosal Trypanosoma cruzi infection Christopher S EickhoffI; Olivia K GiddingsI; Nobuko YoshidaIII; Daniel F HoftI, II, + IDepartment
of Internal Medicine + Corresponding author: hoftdf@slu.edu Received 6 January 2010 Code Number: oc10116 ABSTRACT The potential use of the Trypanosoma cruzi metacyclic trypomastigote (MT) stage-specific molecule glycoprotein-82 (gp82) as a vaccine target has not been fully explored. We show that the opsonization of T. cruzi MT with gp82-specific antibody prior to mucosal challenge significantly reduces parasite infectivity. In addition, we investigated the immune responses as well as the systemic and mucosal protective immunity induced by intranasal CpG-adjuvanted gp82 vaccination. Spleen cells from mice immunized with CpG-gp82 proliferated and secreted IFN-γ in a dose-dependent manner in response to in vitro stimulation with gp82 and parasite lysate. More importantly, these CpG-gp82-immunized mice were significantly protected from a biologically relevant oral parasite challenge. Key words: Trypanosoma cruzi - mucosal immunity - vaccines Trypanosoma cruzi, the causative agent of Chagas disease, infects more than 11 million people and kills an estimated 50,000 individuals annually (CDC 2007). Up to 30% of infected individuals will develop complications from Chagas disease, which include cardiomyopathy and the mega-syndromes (megaesophagus and megacolon). In humans, natural infection occurs primarily through contact with T. cruzi-contaminated reduviid excreta, usually through breaks in the skin or mucosal surfaces. Recent reports of large-scale outbreaks following food or drink contamination demonstrate the dangers of oral T. cruzi infection (Dias et al. 2008, Beltrão et al. 2009, Nóbrega et al. 2009, Pereira et al. 2009, Alarcón de Noya et al. 2010). Because parasite proteins are expressed differentially in distinct T. cruzi life stages, it may be difficult to develop a vaccine that establishes sterile immunity. Glycoprotein-82 (gp82), a stage-specific protein present on the surface of T. cruzi metacyclic trypomastigotes (MT), has been shown to bind to gastric mucin and to facilitate the Ca2+-signaling activity necessary for parasite internalization (Yoshida 2006, 2009). Isolates of T. cruzi that lack gp82 expression are much less efficient at infecting mice orally than those that express high levels of gp82 (Cortez et al. 2003). Antibodies that bind to gp82 inhibit the in vitro infection of epithelial cells (Neira et al. 2003). Although gp82 is only expressed by MT, it could be a good target for immune responses aimed at providing protection against initial immune invasion and intracellular replication cycles. It is well documented that type 1 immune responses are critical for protection against both mucosal and systemic T. cruzi infection (Hoft et al. 2000, Hoft & Eickhoff 2005, Rodrigues et al. 2009). The CpG motifs present within ssDNA are known to induce type 1 immune responses mediated by toll-like receptor 9 (TLR9) stimulation and have been used as adjuvants safely and effectively in mice and humans (Krieg 2000, Dumais et al. 2002, Cooper et al. 2005). Several independent groups have shown the induction of protective immune responses against lethal T. cruzi challenge using CpG mixed with whole T. cruzi lysate or various recombinant T. cruzi proteins (Frank et al. 2003, Araújo et al. 2005, Hoft et al. 2007). In this study, we tested a mucosal T. cruzi vaccine containing the gp82 protein. We show that intranasal vaccination with CpG + gp82 induces protective immune responses against mucosal T. cruzi challenge. MATERIALS AND METHODS Parasites and mice - Female Harlan Sprague Dawley BALB/c mice aged 6-8 weeks were used throughout these studies and housed in Association for Assessment and Accreditation of Laboratory Animal Care International (AAALAC) accredited facilities. Tulahuén strain T. cruzi insect-derived MT (IMT), culture-derived MT and blood form trypomastigotes (BFT) were prepared as previously described (Hoft & Eickhoff 2005, Giddings et al. 2006). Opsonization and T. cruzi challenges - IMT mixed 1:1 with 0.5 mg/mL MAb-3F6 (gp82-specific monoclonal antibody) or isotype control (Sigma, St. Louis, MO, USA) were incubated at RT for 30 min prior to conjunctival or oral challenges, which were performed as previously described (Hoft 1996, Giddings et al. 2006). Immunization with CpG-gp82 - Recombinant gp82 and control glutathione S-transferase (GST) proteins were prepared as previously described (Santori et al. 1996). Oligodeoxynucleotide (ODN) # 1826 (5'-TCCATGACGTTCCTGACGTT-3') containing CpG motifs (underlined) and control ODN were purchased from Coley Pharmaceuticals (Wellesley, MA). BALB/c mice were immunized intranasally twice, two weeks apart, using 10 μg CpG 1826 mixed with 2-10 μg gp82 (or control GST). Measurement of vaccine induced immunity - Four weeks after immunization, spleen cells from vaccinated mice were stimulated at 1 x 106 cells/mL in round-bottom 96-well plates with 0.4-2.0 μg/mL recombinant gp82 or control GST protein. Spleen cells (4 x 106/mL) from these same immunized mice were stimulated in 24-well plates with medium or 10 μg/mL T. cruzi lysate (Hoft & Eickhoff 2005). Three days later, proliferation and IFN-γ secretion were assessed as previously described (Hoft & Eickhoff 2005). Determination of parasite infectivity - Four weeks after the final immunization, some mice (n = 5/group) were challenged with 5,000 BFT subcutaneously and survival was monitored as a means of analyzing systemic protective immunity. Other groups of mice (n = 5-8/group) were challenged orally with 1,000 IMT and parasite replication in mucosal tissues was determined using T. cruzi-specific real-time polymerase chain reaction (PCR). Mice were harvested 11-13 days after mucosal T. cruzi challenge and the levels of parasite replication were measured in draining lymph node and tissue DNA samples using T. cruzi-specific real-time PCR as previously described (Giddings et al. 2006, Hoft et al. 2007). Briefly, DNA was purified from splenic and gastric lymph node tissue (after oral MT challenge) and from submandibular lymph nodes (after conjunctival MT challenge) using a commercially available kit (DNeasy, QIAGEN). T. cruzi-specific real-time PCR was performed using 100 ng of purified DNA per reaction. Ethics - All animal studies were performed in AAALAC accredited facilities (NIH Assurance A3225-01). In addition, all mouse studies were approved by the Institutional Animal Care and Use Committee/Animal Care Committee at Saint Louis University. RESULTS Gp82-specific antibodies reduce T. cruzi infectivity - Our group has shown that the opsonization of T. cruzi MT with faecal extracts containing T. cruzi-specific secretory IgA can reduce parasite infectivity after conjunctival challenge (Giddings et al. 2006). In the present study, we incubated Tulahuén strain IMT with a control or gp82-specific MAb-3F6. The opsonized parasites were then placed on the conjunctiva of anesthetized mice or fed orally to naïve mice. Two weeks later, mice were sacrificed and parasite replication was determined by real-time PCR in tissues taken from sites of early infection. In mice challenged conjunctivally with anti-gp82-opsonized parasites, significantly reduced parasite DNA was detectible in the lymph nodes that drain the site of initial invasion in the nasal cavity relative to mice infected with control opsonized parasites (p < 0.04 by Mann-Whitney U test) (Fig. 1A). Similarly, mice challenged orally with anti-gp82-opsonized parasites had ~4-fold lower amounts of T. cruzi DNA recoverable at the site of initial invasion in gastric tissue compared with controls (Fig. 1B). These results show that gp82-specific antibodies can reduce infectivity at the initial sites of parasite mucosal invasion after both conjunctival and oral T. cruzi challenges. In addition, significantly decreased amounts of parasite DNA were detected in splenic DNA samples isolated from mice challenged orally with anti-gp82 opsonized MT compared with mice challenged with control opsonized MT (p < 0.02 by Mann-Whitney U test) (Fig. 1C). Thus, the systemic spread of parasite infection can be curtailed by opsonization with gp82-specific antibody, suggesting that disseminated disease from T. cruzi can be limited by the neutralization of initial parasite infectivity. Intranasal vaccination with CpG-gp82 induces T. cruzi-specific type 1 immune responses - Systemic immunization with alum + gp82 has previously been shown to induce immune responses that were capable of reducing parasitaemia after systemic MT challenge (Santori et al. 1996). In the present study, we intranazally vaccinated mice with gp82 adjuvanted with CpG, a TLR-9 agonist. Four weeks following the second and final immunization, spleen cells from immunized animals were harvested and stimulated with antigen. The spleen cells from mice immunized with CpG-gp82 proliferated in a dose-dependant manner in response to recombinant gp82 antigen stimulation in vitro (Fig. 2A). Spleen cells from these same vaccinated animals produced IFN-γ upon in vitro stimulation with gp82 (Fig. 2B). Additionally, spleen cells from mice immunized with CpG-gp82 produced IFN-γ after in vitro stimulation with whole T. cruzi lysate (Fig. 2C). The production of IFN-γ by spleen cells from gp82-immunized mice after stimulation with T. cruzi antigens is especially important because type 1 immune responses have been shown to be critical in protecting against both systemic and mucosal T. cruzi challenges (Hoft & Eickhoff 2002, 2005). Intranasal CpG-gp82 isimmunization fails to protect mice against systemic BFT challenge - In order to assess protective immunity induced by CpG-gp82 immunization, immunized mice were challenged systemically or mucosally four weeks after their second and final vaccination. For systemic survival studies, 5,000 BFT diluted in 100 μL PBS were injected subcutaneously at the base of the tail. As seen in Fig. 3A, mice immunized with CpG-gp82 failed to survive lethal T. cruzi challenge and time to death was not prolonged. CpG-gp82 immunization results in protection after a biologically relevant mucosal MT challenge - In order to study mucosal protection, immunized mice were challenged orally with 1,000 IMT. DNA was extracted from stomachs 11-13 days after oral challenge and infection levels were measured using T. cruzi-specific real-time PCR. The gastric DNA obtained from CpG-gp82-vaccinated mice contained significantly reduced amounts of T. cruzi DNA compared with gastric DNA samples from CpG-negative control GST-immunized mice (p < 0.05 by Mann-Whitney U test) (Fig. 3B). DISCUSSION All four T. cruzi life stages have been studied for gp82 expression by simple IFA, Western blot and, more recently, by proteome analysis (Teixeira & Yoshida 1986, Atwood et al. 2005, CDC 2007, The T. cruzi proteome database 2007). It is clear that gp82 is only expressed by T. cruzi MT and not by T. cruzi intracellular amastigotes (AMA) or BFT. The critical role of gp82 in parasite mucosal epithelial invasion makes this T. cruzi glycoprotein an ideal target for mucosal immune responses. It is a highly conserved molecule present in nearly all T. cruzi isolates, including strains Y, CL, F, Tulahuén, G and others (Yoshida 2006). Epimastigotes do not express gp82 and are non-infectious (Manque et al. 2003). In addition, gp82-deficient MT are defective in mucosal infectivity in mice after oral delivery (Cortez et al. 2003). The binding of gp82 to gastric mucin induces intracellular Ca+ increases dependent on ATP consumption, an increase which appears to be required for parasite internalization. The interaction and binding of gp82 to gastric mucin may explain earlier findings in which MT, but not BFT, efficiently infected mice after oral challenge (Hoft 1996). Previous work has shown that antibodies directed against gp82 significantly reduce parasite infectivity both in vitro and after oral in vivo challenges. We have extended these findings by (i) showing that anti-gp82 Abs can block T. cruzi infection via both conjunctival and oral routes and (ii) demonstrating that a mucosal vaccine can significantly reduce mucosal infection after oral T. cruzi challenge. It seems likely that the gp82 gastric mucin-binding epitope overlaps with the gp82-specific MAb-3F6 epitope. Alternatively, immune responses directed against gp82 could block the induction of Ca+/ATP-mediated membrane repair responses, thereby preventing active parasite internalization. It has been reported that mice challenged systemically with MT after gp82 vaccination develop lower parasitaemia compared with control immunized mice (Santori et al. 1996). However, those data were generated using an MT challenge as opposed to a BFT challenge and the endpoint analysis was peak parasitaemia rather than death. In our study, we found no evidence of protection against systemic BFT challenge. Stage-specific gp82 expression on MT (not AMA or BFT) is the most logical explanation for these apparently conflicting data because immune responses directed towards MT may not be effective when BFT not expressing gp82 are used for parasite challenge. Because Santori et al. (1996) used an MT challenge, the gp82-specific immunity that was induced could have partially protected against this initial infecting stage. However, we challenged systemically with BFT that did not express gp82 and, therefore, the gp82-specific immunity is likely to have been irrelevant in recognizing this infection. Many studies investigating T. cruzi vaccine development have focused on the induction of systemic immune responses and have utilized lethal, but perhaps biologically irrelevant, challenge models. As new natural infections in humans occur through contact with IMT-contaminated reduviid excreta, we must investigate the effects of vaccine-induced immunity on these types of infections. Our results showing gp82 vaccine-induced mucosal protection after a biologically relevant IMT challenge are one such example of these types of studies. Future T. cruzi vaccines should encode antigens that can induce both mucosal and systemic immunity. The gp82 antigen could facilitate the induction of optimal mucosal protection, but other antigens will be required for the induction of optimal systemic protection. The prevalence of human T. cruzi infection increases dramatically with the presence of infected dogs in households (Cohen & Gürtler 2001). The removal of infected dog reservoirs alone can reduce the threat of human T. cruzi transmission. Previous studies on dogs immunized with live-attenuated T. cruzi epimastigotes provided partial protection against natural T. cruzi infection (Basombrio et al. 1993). Currently, it may be more feasible to develop a vaccine aimed at preventing systemic and mucosal infection of domesticated animals rather than humans. Our data clearly show that immunization with gp82 induces significant mucosal protection and could be useful in vaccines designed to prevent mucosal infection in dogs. Other vaccines composed of T. cruzi proteins expressed by BFT and/or AMA (including TS and ASP-2) induce strong T-cell responses and systemic protection in mice (Fujimura et al. 2001, Araújo et al. 2005). A vaccine composed of several stage-specific proteins could perhaps lead to optimal protective immunity by inducing immune responses that are relevant for both mucosal and systemic protection. Focusing future vaccine strategies on domestic animals would be much safer than vaccinating humans and could interrupt transmission in households. Thus, a multi-component vaccine that includes gp82 and is aimed at preventing T. cruzi infection in dogs should be explored. ACKNOWLEDGEMENTS To Vanessa D Atayde, for preparation and purification of recombinant gp82. REFERENCES
Copyright © 2010 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc10116f3.jpg] [oc10116f1.jpg] [oc10116f2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}