 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 105, No. 6, 2010, pp. 736-745 ARTICLES Low and high-dose intradermal infection with Leishmania major and Leishmania amazonensis in C57BL/6 mice Denise Fonseca CôrtesI; Matheus Batista Heitor CarneiroI; Liliane Martins SantosI; Talita Correia de Oliveira SouzaI; Tatiane Uceli MaioliI; Ana Luiza C DuzII; Maria Letícia Ramos-JorgeIII; Luis Carlos Crocco AfonsoIV; Claudia CarneiroII; Leda Quercia VieiraI, IV, + IDepartamento
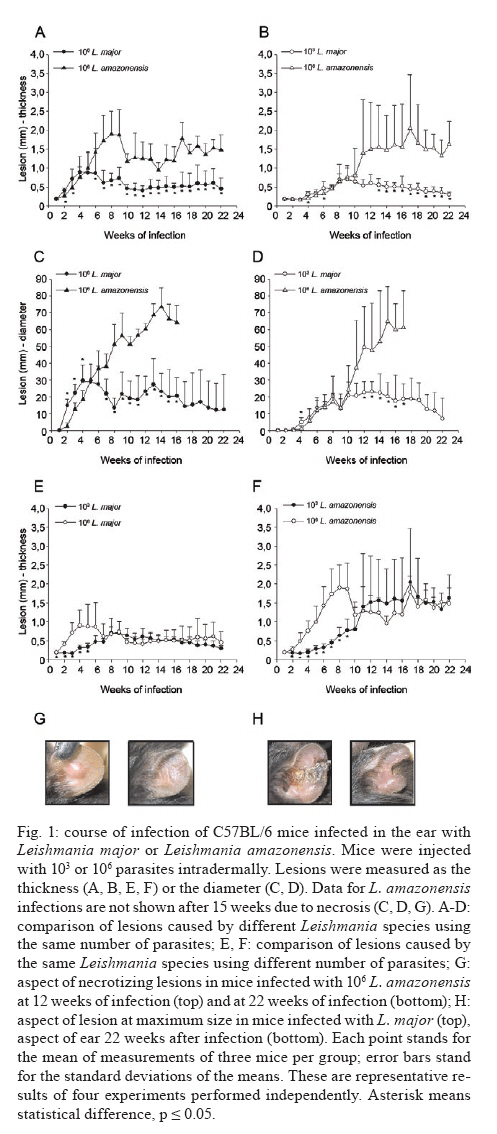
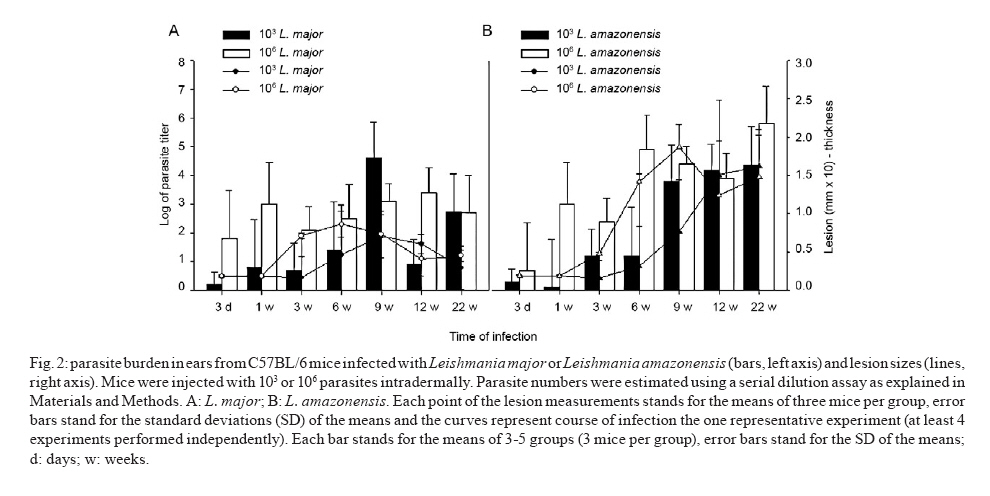
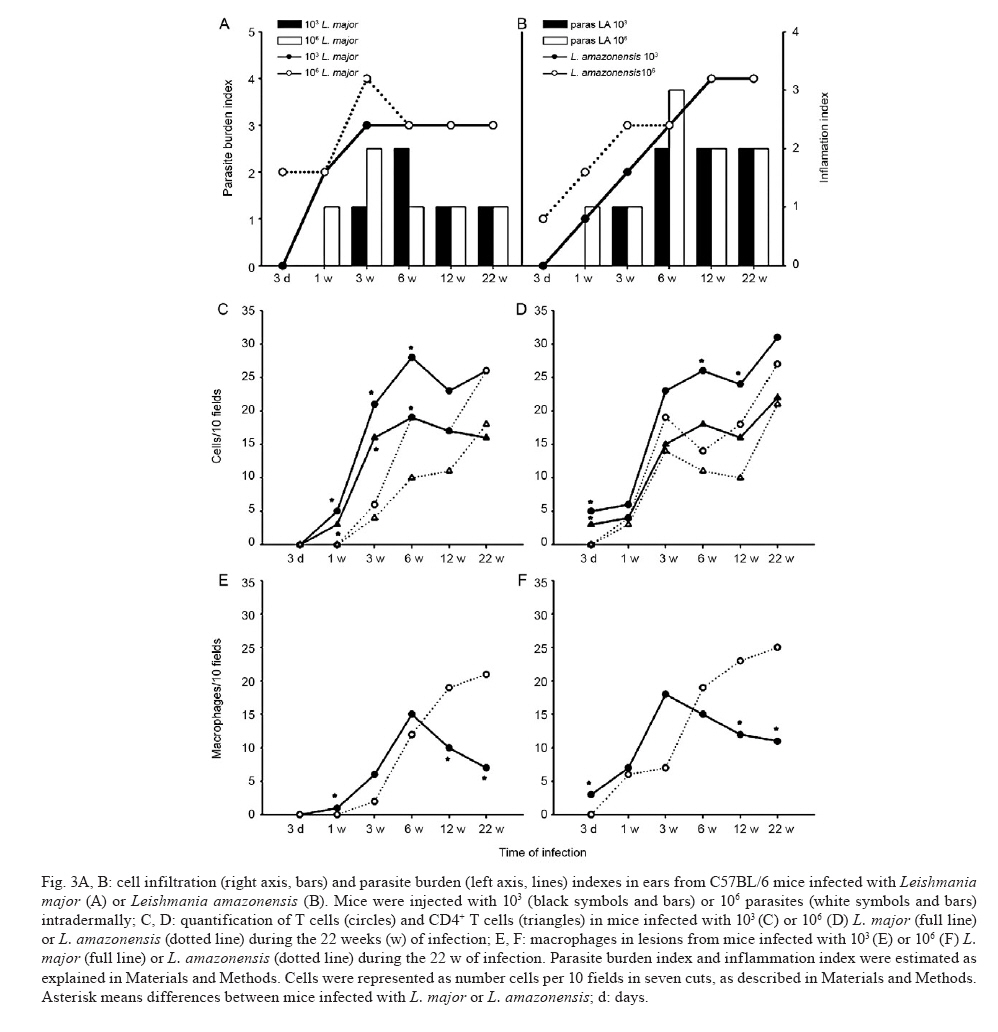
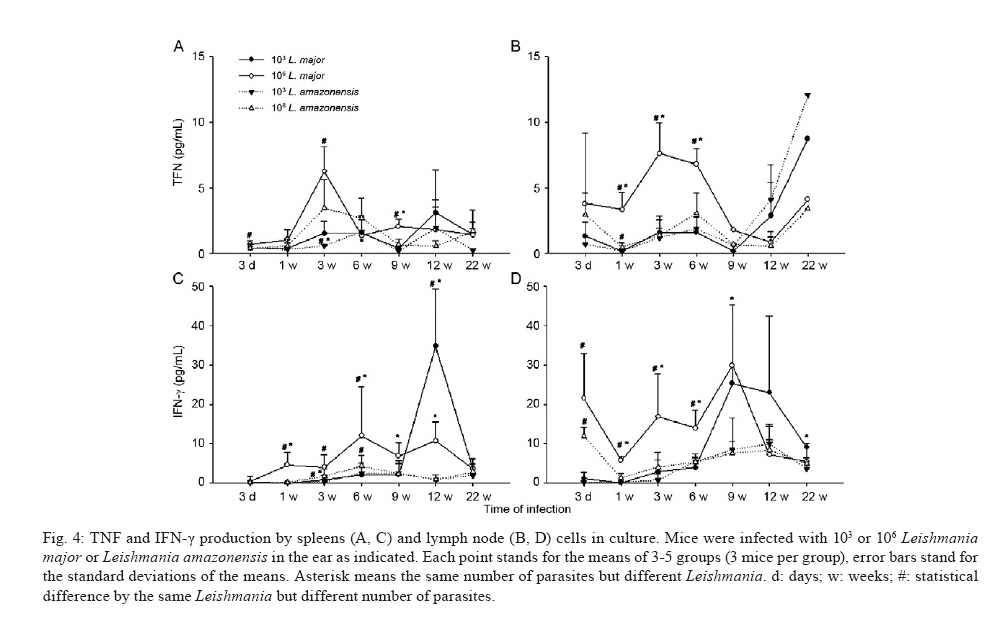
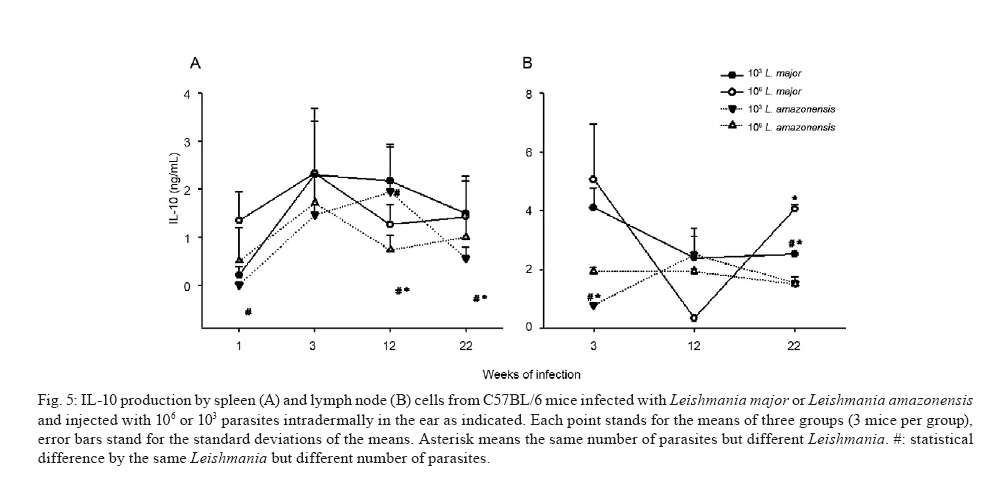
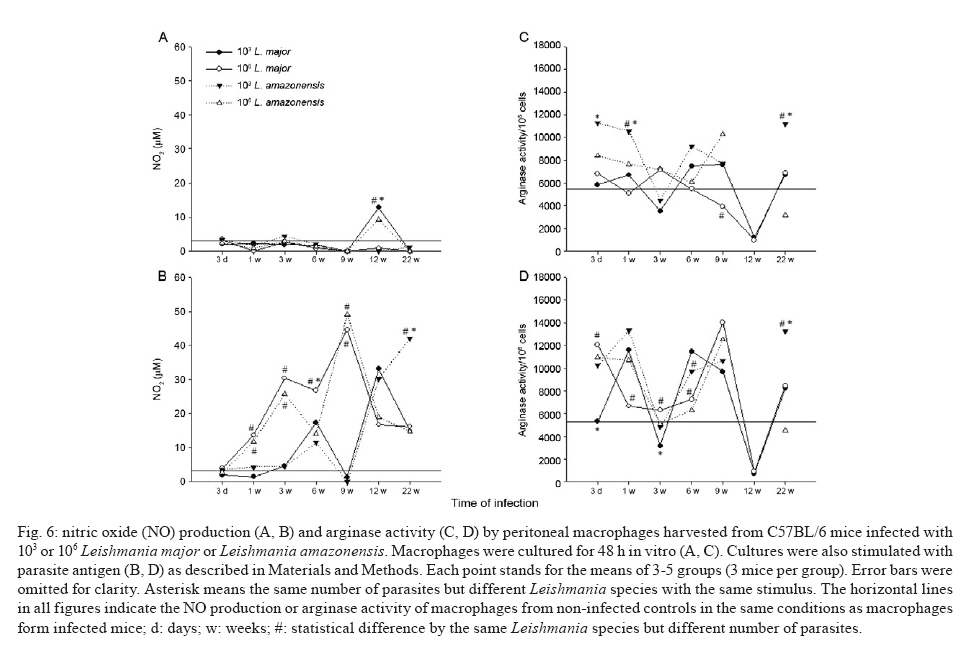
de Bioquímica e Imunologia, Instituto de Ciências Biológicas + Corresponding author: lqvieira@icb.ufmg.br Received 18 December 2009 Financial support: FAPEMIG (CBB87, EDT-538/05), CNPq (350567/1995-6) Code Number: oc10125 ABSTRACT A model of skin infection with Leishmania amazonensiswith low doses of parasites is compared to infection with high doses of L. amazonensis and low and high doses of Leishmania major. C57BL/6 mice were infected with 103 or 106 parasites in the ear and the outcome of infection was assessed. The appearance of lesions in mice infected with 103 parasites was delayed compared to mice infected with 106 Leishmania and parasites were detectable at the infection site before lesions became apparent. Mice infected with L. amazonensisdisplayed persistent lesions, whereas infection with L. major spontaneously healed in all groups, although lymphocytes persisted at the site of infection after healing. Macrophages persisted only in L. amazonensis-infected mice. High-dose L. amazonensis-infected mice produced lower levels of IFN-γ and TNF than mice infected with L. major. No correlation between the persistence of parasites and IL-10 levels and the production of nitric oxide or urea by macrophages was found. We conclude that infection with low doses of L. amazonensisin the dermis changes the course of infection by delaying the appearance of lesions. However, low-dose infection does not change the outcomes of susceptibility and cytokine production described for subcutaneous infection with high numbers of parasites. Key words: Leishmania - L. amazonensis - L. major - infection - cytokines Leishmania amazonensis, like other parasites of the genus, alternates its life cycle between a Phlebotomidae fly and a vertebrate host. L. amazonensis replicates in the gut of flies of the genus Lutzomia and there it differentiates into a metacyclic, infective parasite (Lainson et al. 1987). Infective parasites express different carbohydrates on their surface than do non-infective forms (Saraiva et al. 1986, Courret et al. 1999). Parasites reach the vertebrate host when the fly feeds on the host's blood and regurgitates parasites present in the mouth or adjacent posterior regions (Killick-Kendrick 1990). Once in the vertebrate host, Leishmania metacyclic promastigotes interact with macrophages through ligand receptor-mediated residues on their surfaces and mannose receptors on the macrophage surface (Mosser et al. 1987, 1992, Da Silva et al. 1989). Metacyclic promastigotes interact with macrophages differently than non-infective forms (Courret et al. 1999, Saraiva et al. 2005) and they have been shown to impair the capacity of macrophages to present antigens (Courret et al. 1999). However, most studies use stationary phase cultures, which contain mostly (80%) non-infective forms, for experimental infections (Scott 1989, da Costa et al. 1992, Afonso & Scott 1993, Santiago et al. 1999, Ji et al. 2003). In general, experimental infections with Leishmania are performed using large numbers of stationary phase parasites (105-107) injected subcutaneously (Scott 1989, da Costa et al. 1992, Afonso & Scott 1993, Santiago et al. 1999, Ji et al. 2003). Stationary phase cultures of L. amazonensis contain a mixture of procyclic and metacyclic promastigotes (Courret et al. 1999). Hence, when injected with high numbers of stationary phase parasites, mice are exposed to both infective and non-infective forms. The consequent high levels of antigens and the outcome of infection may be influenced by these protocols. A more natural model has been described for Leishmania major (Belkaid et al. 2000). In this model, intradermal inoculation of low doses of infective metacyclic forms of the parasites revealed that L. major grows at the site of infection without causing a significant lesion; cellular infiltration and lesion development are subsequent to this initial "silent" phase (Belkaid et al. 2000). Intradermal infections with high doses of stationary-phase L. amazonensis promastigotes have been performed previously (Mortatti & Henriques 1990, Courret et al. 2003, Norsworthy et al. 2004, Felizardo et al. 2007), but only two of these studies used low doses of the metacyclic forms of this parasite for infection (Courret et al. 2003, Felizardo et al. 2007). No studies that describe the infection of C57BL/6 mice with low doses of metacyclic parasites by the intradermal route were found. The particular importance of the present study is highlighted by the fact that this model is more similar to the human chronic localized cutaneous leishmaniasis caused by New World Leishmania, which seldom visceralises (Weigle & Saravia 1996). Since the low-dose model for infection with L. major is well-established, we compared the course of infection with low doses of metacyclic L. major and L. amazonensis by the intradermal route in this paper. In addition, the quality of the cellular infiltrate, cytokines produced by lymph nodes and spleen cells and macrophage activity were assessed. MATERIALS AND METHODS Mice - C57BL/6 mice were obtained from the Bioterism Center, Institute of Biological Sciences, Federal University of Minas Gerais. Mice were kept in conventional conditions with barriers, a controlled light cycle and a controlled temperature. Food and water were provided ad libitum. This project was approved by the local ethical committee under the protocol CETEA 040/04. Parasites, infections and antigens - L. amazonensis (IFLA/BR/67/PH8) and L. major (MHOM/IL/80/Friedlin) were maintained in Grace's medium as previously described (Oliveira et al. 2000). Metacyclic promastigotes were purified in a ficoll gradient (Späth & Beverley 2001), washed, suspended in phosphate buffered saline (PBS) (pH 7.3) and counted. Inocula of 103 parasites/10 μL of PBS or 106 parasites/10 μL of PBS were injected in both mouse ears. Lesion development was followed by measuring the diameter and thickness of the lesions using a digital micrometre (Starrett 727, Itu, SP, Brazil). Antigens were prepared from log phase promastigotes that were washed in PBS and submitted to seven cycles of freezing in liquid nitrogen and thawing (37ºC). Suspensions were adjusted to a final concentration of 1 mg of protein/mL and kept at -70ºC until use. Protein concentration was assessed by the Lowry assay (Lowry et al. 1951). Quantification of parasites in ears - Mice were sacrificed by decapitation. Ears were removed and disinfected in 70% ethanol for 5 min and air dried in the laminar flow hood; the two halves were then separated. The dermal side was placed in Roswell Park Memorial Institute medium (RPMI) (GIBCO BRL, Grand Island, NY, USA) containing 100 U/mL penicillin, 100 μg/mL streptomycin (GIBCO BRL) and 125 U/mL collagenase A (Sigma-Aldrich, Inc, St. Louis, MO, USA) for 2 h at 37ºC in a humidified atmosphere containing 5% CO2. The two halves were cut in small pieces and dispersed using a glass tissue grinder. Homogenates were centrifuged at 50 g for 4 min to remove large tissue debris and supernatants were collected and centrifuged at 1,500 g for 15 min. The sediment was resuspended in 0.5 mL of Grace's medium (GIBCO BRL) supplemented with 20% heat-inactivated foetal bovine serum (FBS) (Cultilab, Campinas, SP, Brazil), 100 U/mL penicillin, 100 μg/mL streptomycin and 2 mM l-glutamine (GIBCO BRL). This suspension was serially diluted in a 96-well plate (1:10). Pipette tips were discarded after each dilution. Plates were cultured for 10 days at 25ºC and the last positive dilution was registered as the titre. Results are expressed as the negative logarithm of the titre. Immunohistochemistry - Ear samples were dried and frozen dry at -70ºC. Frozen samples were cut into seven sections of 10 μm, air-dried for 12 h, fixed in cold acetone for 10 min and stored at -70ºC until use. One section was stained with haematoxylin and eosin (H&E). Histological damage was calculated from observation of 10 different fields (40X magnification) of H&E-stained sections from each animal. The histopathological-score grading system used to evaluate the degree of inflammation and parasites was as follows: 0: none; 1: slight; 2: moderate; 3: severe. Other sections (a total of 6) were hydrated with TRIS-buffered saline (TBS) and endogenous peroxidase was blocked by incubation with 1% H2O2 in TBS for 10 min. After washing, the samples were incubated with 5% FBS for 30 min. Then, the sections were washed in 0.05% Tween-20 in TBS for 5 min and later incubated for 1 h with one of the following rat anti-mouse monoclonal primary antibodies diluted in 1% bovine serum albumin-TBS: anti-ε chain of CD3 (T lymphocytes), anti-mouse CD4, anti-mouse CD8, anti-mouse CD40 (B lymphocytes) and anti-mouse Mac3 (macrophage), all of which were from BD Pharmingen (Franklin Lakes, NJ, USA). Controls were incubated only with TBS. The sections were washed three times in TBS for 5 min, incubated with a biotin-conjugated secondary antibody for 30 min in a humidified chamber at room temperature and then incubated with an avidin-biotin-peroxidase complex (ABC kit, Vector, Sydney, Australia). The peroxidase activity was visualized using diaminobenzidine. The samples were counterstained with Harris' haematoxylin, dehydrated and covered with Entellan (Merck, Darmstadt, Germany). The number of positive cells was determined in 10 different fields (100X magnification). Staining for all antibodies used was confirmed by isotype-matched controls. Cell cultures and cytokine assays - Mice were sacrificed at three days as well as one, three, six, nine, 12 and 22 weeks after infection. Cultures of spleen cells and lymph nodes were performed as previously described (Oliveira et al. 2005). Cells were cultured in medium alone or with 50 μg/mL of a homolog antigen for 24 h for TNF assays or for 72 h for IFN-γ and IL-10 assays. IFN-γ was assayed using the monoclonal antibody R46A2, a polyclonal rabbit anti-mouse IFN-γ prepared in our laboratory and an anti-rabbit IgG conjugated with peroxidase (Zymed Laboratories, Inc, San Francisco, CA, USA) (Oliveira et al. 2005). ABTS (Sigma-Aldrich, Inc, St. Louis, MO, USA) and hydrogen peroxide were used as substrates for peroxidase. The detection limit for this assay was 0.03 ng/mL. TNF was assayed using a bioassay (Lattime et al. 1988) with a detection limit of 0.01 pg/mL. Recombinant IFN-γ and TNF-α were used as standards for these assays (R&D Systems Inc, Minneapolis, MN, USA). IL-10 was assayed using a commercially available kit (Peprotech Inc, Princeton Business Park, NJ, USA). The detection limit for IL-10 was 0.03 ng/mL. Macrophages - Macrophages were harvested from mice infected with inocula containing 106 or 103 L. major or L. amazonensis. Mice were injected in the intraperitoneal cavity with 2 mL of 3% thioglycolate broth (Becton, Dickinson and Company, Sparks, MD, USA) at three days or one, three, six, nine, 12 or 22 weeks after infection. Mice were sacrificed three days later and 10 mL of RPMI was injected in the peritoneal cavity to harvest cells. Cells were counted and cultured for 48 h in a culture medium [RPMI containing 10% FBS (Cultilab, Campinas, SP, Brazil)], 100 U/mL penicillin, 100 μg/mL streptomycin and 2 mM l-glutamine (GIBCO BRL). In some experiments, parasite antigen (50 μg/mL), IFN-γ (50 μg/mL) or IL-4 (40 U/mL) was added. Nitric oxide (NO) assay - Supernatants from 48-h macrophage cultures were harvested and NO was evaluated as nitrite by the Griess reaction (Green et al. 1982). Sodium nitrite was used as a standard and the detection limit was 3.9 μM. Arginase assay - Arginase activity in macrophages was assayed as described previously (Corraliza et al. 1994) with a few modifications. Macrophages were obtained as above and cultured for 48 h. After removal of the supernatant, 100 μL of Triton X-100 was added and plates shaken for 30 min. Arginase was activated with 100 μL of MnCl2 (10 mM) in TRIS-HCl (50 mM, pH 7.5) at 55°C for 10 min. Aliquots of samples (5 and 50 μL) were transferred to a fresh 24-well plate with 50 μL of 0.5 mM l-arginine (pH 9.7) and incubated for 60 min at 37°C. The reaction was stopped by the addition of 400 μL of a mixture of H2SO4:H3PO4 :H2O (1:3:7). Subsequently, 25 μL of 9% 1-phenil-1,2-propanodione-2-oxime in ethanol was added and the plates incubated at 95°C for 45 min for color development. Reaction mixtures were read at 540 nm. One unit of enzyme activity is defined as the amount of enzyme that catalyses the formation of 1 μmol of urea/min. A standard curve was performed using urea and the detection limit for the assay was 270 μM urea. Statistical analysis - Data are presented as mean ± standard deviation. The comparison of two groups was analyzed by using a Student's unpaired two-tailed t-test when the distribution of data was found to be normal (Kolmogorov-Smirnov & Shapiro-Wilk tests). When the distribution was not normal, the Mann-Whitney test was used. Differences were considered statistically significant when p d" 0.05. For comparison of more than two groups, a one-way ANOVA was performed. If a significant main effect or association was identified (p d" 0.05), the respective group means were compared using Bonferroni's t-test. RESULTS Course of infection with L. amazonensis in mice infected with low doses of parasites in the skin - C57BL/6 mice were infected in the ear (intradermal inoculum) with 103 or 106 L. major or L. amazonensis and the course of infection was followed over time. We found that mice infected with 106 L. major developed lesions that healed spontaneously, whereas L. amazonensis infection caused a chronic lesion that was not completely uncontrolled but also did not heal (Fig. 1A, C) and started necrotizing from 10 weeks after infection (Fig. 1G, top). The decreased lesion size shown in Fig. 1A corresponded to the onset of necrosis. Several mice suffered loss of tissue at 15 weeks after infection (Fig. 1G, bottom). When mice were infected with 103 L. amazonensis, the same profile of partial susceptibility was seen (Fig. 1B, D). However, the detection of lesions was delayed by several weeks when compared with that in mice infected with high doses of parasites (Fig. 1E, F). Very few mice infected with low doses of L. amazonensis showed necrosis. Mice infected with low doses of L. major developed lesions in ears that peaked at eight weeks (Fig. 1B, D, H top) and slowly healed (Fig. 1H, bottom). Mice infected with low doses of L. amazonensisdeveloped lesions equivalent in size to those of L. major-infected mice, but these lesions did not show signs of healing up to 22 weeks after infection. Quantification of parasites at the site of infection - Parasites were quantified at the site of infection at several time points. Parasite growth preceded the detection of significant lesions in mice infected with low doses of L. major (Fig. 2A). High numbers of parasites were also present at the site of infection with 106 L. major at three days and one week after infection. Lesions were detected in this group three weeks after infection (Fig. 2A). With regard to infection with L. amazonensis, Fig. 2B demonstrates that a high number of parasites were found one week after infection with the high-dose. Lesions were detectable three weeks after infection (Fig. 2B). When mice were infected with a low-dose of parasites, the detection of lesions was delayed. Three weeks after infection, significant growth of parasites compared to the growth at week one was found; the number of mice bearing lesions was almost insignificant. Nine weeks after infection, however, groups infected with high and low doses of L. amazonensis had similar numbers of parasites, even though the number of mice bearing lesions from mice infected with 103 parasites equaled that from mice infected with 106 parasites only 12 weeks after infection. There was a positive correlation between the number of parasites and lesion size for mice infected with L. amazonensis (r2 = 0.972 and 0.923 for the low and high doses, respectively), even though lesions appeared after parasite growth. For both L. amazonensis and L. major, mice infected with high numbers of infective parasites show more parasites at the site of infection at early time points. However, the total number of parasites at the time of peak lesion size was similar, regardless of the initial parasite inoculum. Cellular infiltration - Inflammation and parasite burden indices were established to quantify the aspects of lesion histology described above. Parasite growth occurred prior to the detection of lesions in low-dose infected mice (Fig 2, 3A, B). Moreover, cellular infiltration was delayed in low-dose infected mice and correlated well with the delayed detection of lesions. Although lesions were practically non-existent at the end of the course of L. major infection (Fig. 1E, F, H), it was interesting to notice that some cellular infiltration and parasites were present at the site of infection. Immunohistochemistry was performed in histological cuts to determine the nature of the cellular infiltrate found in each of the groups. As shown in Fig. 3 C-F, the inflammatory infiltrate consisted of CD4+ T cells, macrophages and lower numbers of CD8+ T and B cells (not shown). In mice infected with L. amazonensis, CD4+ T cells appeared at earlier time points after infection with 106 parasites (Fig. 3B). CD4+ T cells were present at similar levels in mice infected with high and low doses of L. major at the time-points examined (Fig. 3D). CD8+ T cells were present at lower numbers than CD4+ T cells and followed similar kinetics. B cells were also found in very small numbers and their increase over the course of infection was not always evident (data not shown). Higher numbers of macrophages infiltrated into the site of infection in mice infected with 106 parasites (Fig. 3E, F) and this infiltration was somewhat delayed in mice infected with 106 L. amazonensis compared with mice infected with 106 L. major (Fig. 3F). Regardless of the size of the parasite inoculum, macrophages were present at lower numbers at 12 and 22 weeks in mice infected with L. major than in mice infected with L. amazonensis (Fig. 3E, F). CD4+ T cells, on the other hand, were present at high levels in all four groups of mice at the later times after infection (Fig. 3C, D). Cytokine production by lymph node and spleen cells - Cytokines were measured over the course of infection with high or low doses of L. major and L. amazonensis. Results for TNF and IFN-γ are shown in Fig. 4. Cells from spleens and lymph nodes of mice infected with the high-dose of L. major showed greater production of TNF in vitro than cells from mice infected with 106 L. amazonensis for most time points (Fig. 4A, B). The same was not true when inocula containing 103 parasites were compared. In addition, spleen cells from mice infected with 106 parasites produced more TNF than spleen cells from mice infected with 103 parasites at most time points when the same Leishmania species was considered. Interestingly, all groups showed an increase in TNF production by lymph node cells at later times after infection (Fig. 4B). Mice infected intradermally with 106 L. major produced higher levels of IFN-γ than mice infected with the same dose of L. amazonensis (Fig. 4C, D). For the low-dose of parasites, this difference was not found. Cells from mice infected with a high-dose of L. major produced more IFN-γ than cells from mice infected with a low-dose of parasites. Interestingly, at nine weeks, we found that a larger amount of IFN-γ was produced by cells from both the spleen and lymph nodes in the low-dose group (Fig. 4C, D). IFN-γ production did not differ significantly between cells from low and high-dose L. amazonensis-infected mice, despite the lower number of parasites in mice infected with the low dose (Fig. 2B, from 3 days to 9 weeks). We also investigated the production of IL-10 by cells from spleens and lymph nodes in culture. Results are shown in Fig. 5 and no clear-cut differences were found between L. major and L. amazonensis or mice infected with low and high doses. NO production and arginase activity by macrophages from mice infected with high and low doses - Two pathways for l-arginine metabolism were investigated. Macrophages from the peritoneal cavity of mice infected with high or low doses of L. major or L. amazonensis were harvested at several time points and NO production in culture supernatants as well as arginase activity in cells were determined. Macrophages from infected mice did not present spontaneous production of NO (Fig. 6A). However, when stimulated with the parasite antigen in vitro, we found that there was generally a greater production of NO by macrophages from mice infected with the high-dose of parasites up to nine weeks after infection (Fig. 6B). In addition, there were no outstanding differences in NO production by macrophages from mice infected with L. amazonensis or L. major at the same dose. When we added IFN-γ to the macrophage cultures, background production (by non-infected mice) was very high; no differences were found among groups (data not shown). Arginase activity by peritoneal macrophages from infected and non-infected mice was very similar (Fig. 6C). Three days, one week and 22 weeks after infection, spontaneous arginase activity was higher in macrophages from mice infected with 103 L. amazonensis than 103 L. major. When parasite antigens were added, there was a general tendency toward decreased activity at three and six weeks after infection (Fig. 6D). However, no clear correlations among levels of arginase, numbers of parasites and parasite species were found. Addition of IFN-γ or IL-4 did not alter the arginase production found in cultures to which the antigen as added (data not shown). DISCUSSION A natural model of infection with L. major was recently described (Belkaid et al. 2000). In this model, a low-dose of infective metacyclic promastigotes (102-103) was inoculated intradermally in mice. This approach revealed several aspects of the infection that were otherwise masked by the traditional high doses of stationary phase parasites (106-107), which contain non-infective forms, via the subcutaneous route in the footpad. Hence, we decided to apply the same model to infections with L. amazonensis, a chronic infection (Afonso & Scott 1993) that simulates chronic infection in humans (Weigle & Saravia 1996). We used purified metacyclic parasites, assuring that all parasites injected were infective. Overall, we found that infection with a low-dose of L. amazonensis in the ear followed the pattern of infections with a high-dose in the footpads: non-healing lesions persisted for several weeks, parasites were present in these lesions in high numbers and low levels of IFN-γ and TNF were produced in response to this parasite. Mice infected with L. amazonensis depicted lesions similar to those from mice infected with the same dose of L. major up to the peak of lesion size with L. major. From that point on, lesions in L. major-infected mice proceeded to heal; in contrast, lesions in L. amazonensis-infected mice did not. These findings are in accordance with data previously published for high doses of parasites injected subcutaneously (Afonso & Scott 1993, Soong et al. 1996, Ji et al. 2003). We also show that, similarly to L. major (Belkaid et al. 2000), L. amazonensis grows silently at the site of infection prior to the appearance of lesions. Similarly to the L. major model, this was especially evident when a low-dose of parasites was used. When low and high-dose lesions are compared, there is an evident delay in the growth of the low-dose lesions. This coincides with the smaller number of parasites and also, perhaps more importantly, with a delay in the appearance of T cells and macrophages. Fewer macrophages were seen at later time points in L. major-infected animals, in accordance with the decreased lesion size in these animals. In contrast, macrophages persisted at high numbers in lesions from L. amazonensis-infected mice. In general, and especially in low-dose infections, the cellular infiltrates were delayed in L. amazonensis-infected mice when compared to L. major-infected mice. This finding is in accordance with the previously described delayed expression of chemokines and cytokines during infection with the former parasite (Ji et al. 2003). Although smaller numbers of parasites were found in mice infected with a low-dose of L. amazonensis, the same number of parasites (on the order of 104-105) eventually persisted in lesions from mice infected with high or low doses. The persistence of macrophages in lesions from L. amazonensis-infected mice would account for the persistence of the parasite, since this is the host cell. Nevertheless, no reason for this persistence was found by us or is described in the literature. As previously noted, chemokine expression is delayed in L. amazonensis-infected mice (Ji et al. 2003). At later time points, chemokine expression in lesions is similar between L. amazonensis and L. major-infected mice; however, these authors only determined cytokine and chemokine expression up to the peak of L. major infection. We tried to determine cytokine and chemokine levels in ears at later time points, but we were unsuccessful. A technical problem yet to be identified prevented us from obtaining good mRNA extractions and reverse-translations. As we have successfully resolved cytokines and chemokines in footpads previously (Santiago et al. 2004, 2006), the reason for our problem is likely related to the ear tissue. IFN-γ production is remarkably low in L. amazonensis-infected mice, especially when production in mice infected with 106 parasites is compared to that in L. major-infected mice. The low IFN-γ production is, in our experience, at least partially responsible for the maintenance of parasites in tissues. Although IFN-γ has been shown to favor growth of L. amazonensis amastigotes in macrophages in vitro and in vivo (Qi et al. 2004), this cytokine seems to participate in the late control of L. amazonensis (Pinheiro & Rossi-Bergmann 2007). Moreover, when IFN-γ production is elevated by vaccination, a small but significant decrease in parasite levels is seen (Vanloubbeeck & Jones 2004, Hernández et al. 2006, Hernández Sanabria et al. 2007, González-Lombana et al. 2008). It is interesting to notice that L. amazonensis fails to trigger a Th2 response in C57BL mice (Afonso & Scott 1993, Jones et al. 2002, Ji et al. 2003, Hernández et al. 2006), even though lesions are quite severe and lead to necrosis and a loss of tissue. IL-10 is not responsible for the lack of parasitism control or resolution of lesions, since there is no significant upregulation of this cytokine when lesions from L. amazonensis and L. major-infected mice start to differ. Moreover, no significant differences in the course of infection with L. amazonensis were found between wild-type and IL-10 knockout mice (Jones et al. 2002, González-Lombana et al. 2008). Accordingly, no increased expression of arginase was found when footpad lesions from mice infected with L. amazonensis or L. major was compared. On the contrary, both NO synthase and arginase were upregulated in footpads of mice infected with L. major compared to those infected with L. amazonensis (Santiago et al. 2006). In fact, the potential for NO and arginase production by macrophages harvested from infected mice at the time points prior to and during the divergence of lesion sizes, which were determined in vitro, were quite similar. It has been previously shown that macrophages from mice infected with L. major produce more TNF than do those from mice infected with L. amazonensis (Gomes et al. 2003). Moreover, CBA mice produce less TNF when infected with L. amazonensis than when infected with L. major (Lemos de Souza et al. 2000). Indeed, when we infected mice with a high-dose of parasites, we found that lymph nodes from mice infected with L. major produced higher levels of TNF than lymph nodes from mice infected with L. amazonensis. However, when low doses of parasites were compared, this differential production of TNF was not found. This finding is in accordance with the similar levels of NO produced by macrophages from mice infected with either parasite. In the context of our investigation, the inability of mice to kill the parasite appears to be due solely to the lower production of IFN-γ. The reason for the diversity of pathogenesis between L. major and L. amazonensis remains unknown. There is very little genetic diversity among the three Leishmania species with sequenced genomes. It has been postulated that the separation of the former continent Gondwana gave rise to two subgenera: Viannia and Leishmania (Tuon et al. 2008). The fact that both L. major and L. amazonensis belong to the same subgenus Leishmania, however, suggests a genetic proximity. It is likely that, as suggested by Lynn and McMaster (2008), a few differences in protein expression might account for the diverse pathology shown by different Leishmania species, especially given the complexity of the immune response and the parasite strategies to escape it (Lynn & McMaster 2008). We have studied the outcome of infection with L. amazonensis using a more natural model than previously reported. We used a lower dose of purified infective parasites introduced via the intradermal route. This method is in opposition to the high doses of stationary phase parasites introduced via the subcutaneous route generally reported in the literature. We showed that the outcome of infection was similar to that previously described (i.e., with non-healing lesions, low production of IFN-γ and no TNF). In addition, we found that there was a phase of silent parasite growth at the beginning of the infection. We showed that macrophages and T cells persisted at the site of infection. The number of parasites at the site of infection became high, regardless of the number of parasites initially used for the inoculation. We believe that this model will be useful to clarify the cause of non-healing lesions due to infection with L. amazonensis. REFERENCES
Copyright © 2010 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc10125f2.jpg] [oc10125f4.jpg] [oc10125f5.jpg] [oc10125f3.jpg] [oc10125f6.jpg] [oc10125f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}