 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 105, No. 6, 2010, pp. 786-791 ARTICLES Expression of bacterial virulence factors and cytokines during in vitro macrophage infection by enteroinvasive Escherichia coli and Shigella flexneri: a comparative study Silvia Y BandoI, II; Ana CR MorenoI; José AT AlbuquerqueI; Juliana MK AmhazI; Carlos A Moreira-FilhoII; Marina B MartinezI, + IDepartamento
de Análises Clínicas e Toxicológicas, Faculdade de Ciências
Farmacêuticas, Faculdade de Medicina, Universidade de São Paulo,
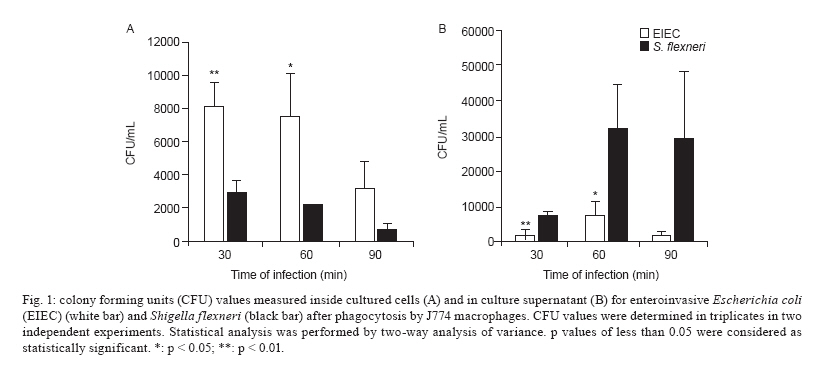
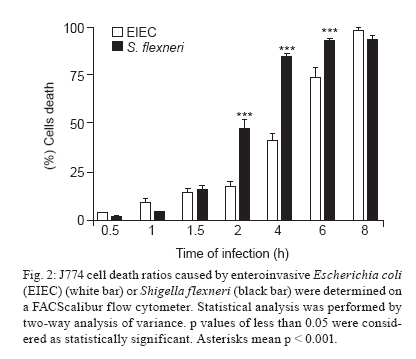
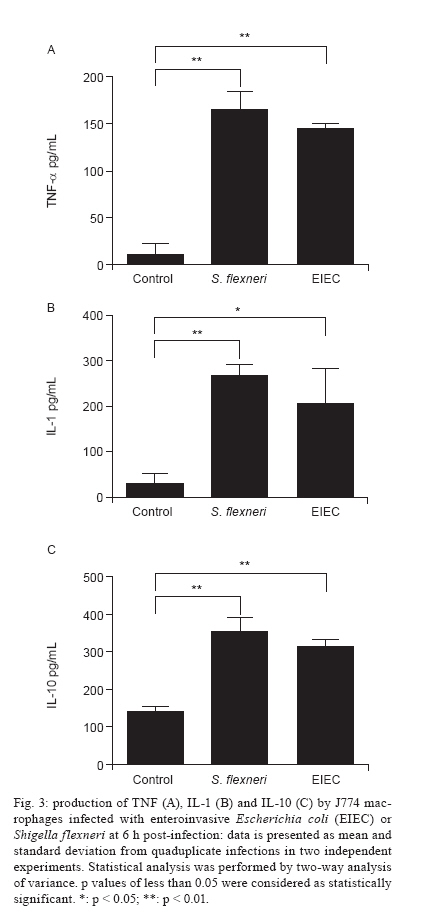
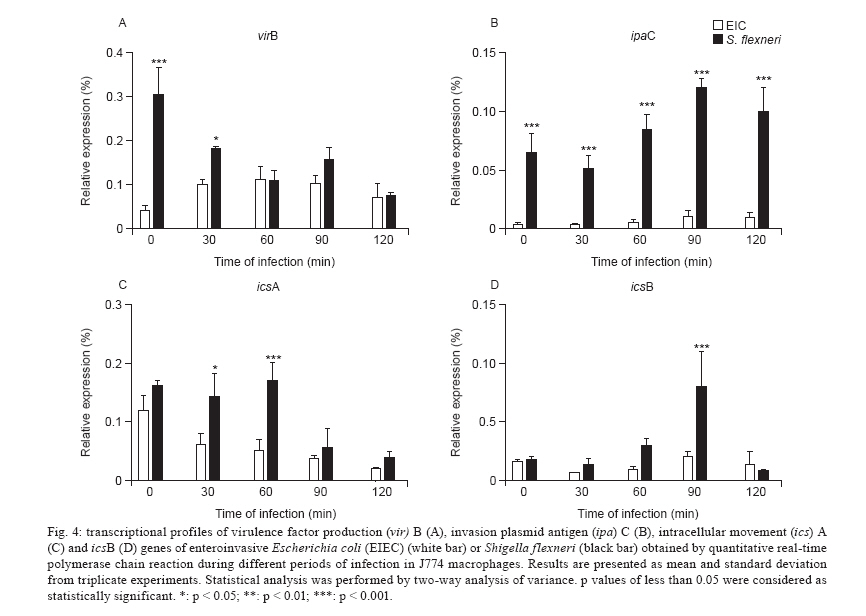
Av. Prof. Lineu Prestes 580, 05508-000 São Paulo, SP, Brasil + Corresponding author: mbmartin@usp.br Received 20 January 2010 Financial support: FAPESP (203/01269-1) Code Number: oc10132 ABSTRACT Enteroinvasive Escherichia coli (EIEC) and Shigellaspp cause bacillary dysentery in humans by invading and multiplying within epithelial cells of the colonic mucosa. Although EIEC and Shigellashare many genetic and biochemical similarities, the illness caused by Shigellais more severe. Thus, genomic and structure-function molecular studies on the biological interactions of these invasive enterobacteria with eukaryotic cells have focused on Shigella rather than EIEC. Here we comparatively studied the interactions of EIEC and of Shigella flexneriwith cultured J774 macrophage-like cells. We evaluated several phenotypes: (i) bacterial escape from macrophages after phagocytosis, (ii) macrophage death induced by EIEC and S. flexneri, (iii) macrophage cytokine expression in response to infection and (iv) expression of plasmidial (pINV) virulence genes. The results showed thatS. flexneri caused macrophage killing earlier and more intensely than EIEC. Both pathogens induced significant macrophage production of TNF, IL-1 and IL-10 after 7 h of infection. Transcription levels of the gene invasion plasmid antigen-C were lower in EIEC than in S. flexneri throughout the course of the infection; this could explain the diminished virulence of EIEC compared to S. flexneri. Key words:EIEC - Shigella - in vitro infection - cytokines - virulence factors Infectious diarrhea is one of the main causes of childhood death, especially in developing countries (Bryce et al. 2005) and Escherichia coli is a major diarrhea-causing pathogen in children under five years of age (Petri et al. 2008, Moreno et al. 2010). E. coli isolated from patients with dysentery was found to cause experimental keratoconjunctivitis in guinea pigs in the early 1960s (Trabulsi et al. 1967, DuPont et al. 1971). These strains, thereafter named enteroinvasive E. coli (EIEC), possess genetic and biochemical characteristics similar to Shigella (Silva et al. 1980, Sansonetti et al. 1982, Kaper et al. 2004). EIEC and Shigella cause bacillary dysentery in humans by invading and multiplying within epithelial cells of the colonic mucosa, resulting in an intense inflammatory response characterized by abscesses and ulcerations (Hale 1998, Parsot et al. 2005). The genes required for bacterial entrance into host cells are harboured in a 230-kb virulence plasmid (VP) present in Shigella and EIEC strains (Harris et al. 1982, Sansonetti et al. 1982, Silva et al. 1982). Transcription of invasion plasmid antigen (ipa) genes is regulated by two VP-encoded proteins: VirF and VirB (Dorman et al. 2001). VirB also controls transcription of the VP genes that code for the proteins of the type three secretion system (TTSS) apparatus. Intracellular movement (ics) A and icsB play a crucial role in bacterial dissemination: the first leads to bacterial invasion and intracellular spreading and the latter aids in escape from autophagy. Shigella translocates through M cells of the intestinal epithelium to invade the epithelial layer by the basolateral pole (Sansonetti 1991, Ménard et al. 1996, Sansonetti et al. 1996). Shigella can survive and multiply inside macrophages, escape from phagocytic vacuoles and induce apoptosis (Sansonetti et al. 1986, Zychlinsky et al. 1992, 1996, Nonaka et al. 2003, Kubota 2006). Apoptosis of infected macrophages is promoted by caspase-1 activation. Caspase-1 cleaves the IL-1 and IL-18 precursors to their mature forms, triggering a mucosal inflammatory response (Navarre & Zychlinsky 2000, Kubota 2006, Suzuki et al. 2007). An inoculum of 102 Shigella cells is sufficient for infection, but as many as 106 EIEC cells are necessary (DuPont et al. 1971), indicating the diminished virulence of EIEC. The cellular processes and genes related to bacterial invasion have been investigated primarily in Shigella flexneri and thus remain poorly understood in EIEC. The interaction of EIEC with macrophage is relevant because bacterial survival inside phagosomes and the subsequent escape from macrophages are critical steps for the establishment of an infection. Therefore, the aim of this study was to compare the expression of genes involved in the regulation of virulence factor production (virF and virB) and cell lysis (ipaB, ipaC) as well as spreading and icsA and icsB. We also measured the cytokine response upon in vitro infection of macrophages with EIEC or S. flexneri. SUBJECTS, MATERIALS AND METHODS Bacterial strains - The M90T strain (invasive S. flexneri 5a) was kindly donated by Philip Sansonetti (Pasteur Institute). EIEC strain FBC124-13, serotype O124:H-, was isolated from diarrheic faeces and was Sereny test-positive (Gibotti et al. 2004). Bacteria were grown at 37ºC for two days on trypticase soy agar plates containing 0.02% Congo Red. Prior to cell infection, a single red colony of each strain was inoculated in trypticase soy broth and grown at 37ºC with agitation (150 rpm) to an OD625 of 0.1, which corresponds to 1 × 108 colony forming units (CFU)/mL. Cell culture and macrophage infection - The J774 macrophage-like cells were maintained in Roswell Park Memorial Institute (RPMI) 1640 medium supplemented with 10% fetal bovine serum (FBS) and penicillin-streptomycin (100 u/mL-100 μg/mL) in a 5% CO2 incubator at 37ºC. Twenty-four hours prior to infection macrophages were plated in 24-well plates at a density of 1 × 106 cells per well. The medium was then replaced by fresh RPMI 1640 containing 10% FBS (without antibiotics) and the macrophages were infected with a multiplicity of infection (MOI) of 10 bacteria per cell, centrifuged at 100 g for 5 min and incubated for different periods of time. Bacterial survival inside the macrophages - Macrophages were infected with S. flexneri or EIEC for zero (cells harvested just after bacterial inoculation), 30, 60 and 90 min. The cells were then incubated with medium containing 50 μg/mL gentamicin for 50 min to kill non-phagocytosed bacteria and to prevent reinfection. The infected macrophages were washed three times with PBS. PBS containing 0.1% Triton X-100 was then added to each well to lyse the macrophages. The recovered bacteria were resuspended in 0.9% NaCl, serial dilutions were plated on MacConkey agar and the CFU were counted following an overnight incubation of the plates at 37ºC. Bacterial escape and macrophage death - To assess bacterial escape after phagocytosis and determine whether this process was accompanied by macrophage death, J774 cells were infected for an hour with EIEC or S. flexneri and then the medium was replaced by fresh RPMI 1640 containing 10% FBS and 50 μg/mL of gentamicin. Following different periods of post-infection incubation (30, 60 or 90 min) in medium without antibiotic, supernatant dilutions were plated to determine the CFU. The macrophages were then resuspended in a hypotonic fluorochrome solution (0.1% sodium citrate, 0.1% Triton X-100 and 50 μg/mL of propidium iodide) and analyzed on a FACScalibur flow cytometer (Becton Dickinson, Mountain View, CA, USA). Detection of cytokines by enzyme-linked immunosorbent assay (ELISA) - The J774 macrophages were infected as described for the bacterial escape assay, however with an MOI of 1:10. After one hour of infection, the medium was replaced by fresh RPMI 1640 containing 10% FBS and 50 μg/mL of gentamicin. Supernatants of post-infected J774 cell cultures were collected after 6 h of incubation. Concentrations of IL-1, TNF and IL-10 were determined by ELISA according to the manufacturer's instructions (Peprotech, Mexico). Expression of EIEC and S. flexneri virulence genes in infected macrophages - Total RNA from infected macrophages was extracted with a RNeasy Mini Kit (Qiagen, Hilden, Germany), as described by the manufacturer. Contaminating genomic DNA was removed using the RNase-free DNase Data Set (Qiagen, Hilden, Germany) and samples were frozen at -80°C until use. Total RNA was quantified by A260 measurements (NanoDrop® ND-1000 Spectrophotometer; Amersham Biosciences) and diluted to 10 ng/μL prior to cDNA synthesis. Bacterial mRNA was reverse transcribed into cDNA using the Superscript II First-Strand Synthesis System with random hexamers, as described by the manufacturer (Invitrogen, Carlsbad, CA, USA). The cDNA was sequenced to check the primer and probe adequacy. Reverse transcriptase polymerase chain reaction (RT-PCR) products were obtained by PCR amplification for 30 cycles as follows: 95°C for 45 s, 50-60ºC (Table I) for 45 s and 72ºC for 2 min, with an initial denaturation step of 95ºC for 5 min and final extension step of 72ºC for 7 min. The virulence genes studied here and their respective primers are listed in Table I. Quantitative real-time PCR (qRT-PCR) analyses were carried out using a TaqMan assay on an ABI Prism 7000 Sequence Detection System version 1.6 (Applied Biosystems). The parameters for PCR were as follows: 50ºC for 2 min, 95ºC for 10 min and 50 cycles of 95ºC for 15 s and 60ºC for 1 min. The primers and the fluorogenic probes (Assays-by-design, Applied Biosystems) used in this study are described in Table II. For each reaction, 16S rRNA was used as an endogenous housekeeping gene control. Data were normalized to control levels and analyzed by the standard curve method. The relative comparison method was used to evaluate the expression levels of the bacterial virulence genes. Statistical analysis - The data were analyzed by a two way analysis of variance and the results were confirmed through multiple comparisons by Tukey's test. Differences between groups were considered significant when p < 0.05. RESULTS Survival and escape of EIEC and S. flexneri during infection of J774 macrophages - The number of viable intracellular EIEC was higher than S. flexneri after 30 and 60 min of infection (Fig. 1A). The amount of bacteria recovered from supernatants confirmed a diminished capacity of EIEC to escape from macrophages. After 30 or 60 min of incubation in gentamicin-free medium, macrophage cultures infected with S. flexneri yielded 4-fold higher CFU in comparison with those infected with EIEC (Fig. 1B). To confirm that these differences were due to escape from macrophages and not to differences in bacterial multiplication rate, growth curves for EIEC and S. flexneri in LB media were obtained and found to be very similar (data not shown). J774 cells death caused by bacterial infection - Macrophage death rates due to S. flexneri and EIEC infections were analyzed throughout 8 h of infection (Fig. 2). We observed 50% cell death in cultures infected with S. flexneri at 2 h post-infection, whereas in EIEC infected cultures we observed only 20% cell death. This difference was proportionally maintained until the 4th h of infection and convergence of cell death ratios was observed after 8 h of infection, indicating that EIEC is less efficient than S. flexneri at macrophage killing. Cytokines expression after J774 infection with EIEC and S. flexneri - We determined the expression levels of IL-1, IL-10 and TNF-α following infection of J774 macrophages with EIEC or S. flexneri to evaluate if the cytokine expression profile differs based on the infecting bacterial species. We observed a similar increase in IL-1, IL-10 and TNF concentrations measured in the supernatants of J774 cells infected with EIEC or S. flexneri compared to supernatants from uninfected cultures (Fig. 3). Differential expression of selected virulence genes in EIEC and S. flexneri during J774 cell infection - qRT-PCR was performed to measure virF, virB, ipaB, ipaC, icsA and icsB transcription rates every 30 min during the 1st 2 h of infection. We observed a very low ipaC transcription level in EIEC, but not in S. flexneri, throughout the course of the infection (Fig. 4). This was the only statistically significant difference between EIEC and S. flexneri virulence gene expression at all time intervals. Higher transcription levels were also observed for virB and icsA in S. flexneri during the initial 30 min and during the 30-60 min interval, respectively. Transcription of icsB was elevated in S. flexneri at 90 min after infection. No significant differences between EIEC and S. flexneri were found in the transcription levels of virF and ipaB (data not shown). The expression of EIEC or S. flexneri virulence genes was also studied when bacteria were cultured in medium without contact with J774 cells and we observed no difference in gene expression between EIEC and S. flexneri (data not shown). DISCUSSION Shigella pathogenicity is determined by several genes encoded by the VP promoting bacterial invasion, escape and proliferation across enterocytes. However, this mechanism remains to be fully elucidated for EIEC. In this work, we aimed to clarify differences between EIEC and S. flexneri regarding macrophage escape mechanisms, expression of associated virulence genes and the macrophage cytokine expression profile induced by these bacteria. Infection with S. flexneri causes rapid macrophage death (Clerc et al. 1987, Nonaka et al. 2003), which constitutes an evasion mechanism from the innate immune system (Kubota 2006, Schroeder & Hilbi 2008). Therefore, the first step in the present investigation consisted of determining whether EIEC and S. flexneri survive in and escape from macrophages in a similar way. EIEC was less efficient in macrophage killing during the initial 4 h of infection than S. flexneri (Figs 1, 2) which may indicate differences in the expression of virulence genes. The virulence gene ipaC is involved in bacterial escape from the phagocytic vacuole and is responsible for lysis of the phagosome membrane (Harrington et al. 2006, Schroeder & Hilbi 2008). This gene is also responsible for human epithelial cell invasion by targeting the TTSS, which leads to the efficient injection of effector proteins that improve the invasion process (Jaumouillé et al. 2008). In the present study, we detected a very low expression of ipaC in EIEC throughout the infection compared to S. flexneri (Fig. 4). EIEC shares almost complete identity of the ipaC nucleotide sequence with S. flexneri (Gibotti et al. 2004), suggesting that an EIEC chromosomal factor(s) could modulate ipaC transcription inside macrophages. However, further studies are necessary to investigate this hypothesis. No differences in the expression of the pro-apoptotic gene ipaB were detected between EIEC and S. flexneri. This gene encodes for a protein that activates caspase-1, which initiates macrophage apoptosis and the cleavage of proinflammatory cytokines (Zychlinsky et al. 1994, Schroeder & Hilbi 2008). Recently, we studied the expression of EIEC and S. flexneri VP genes in a different experimental model, focusing on intestinal cell invasion and dissemination. We demonstrated that the disease triggered by EIEC may be restricted to a definite infection site, meaning that it is not capable of disseminating beyond a certain point to extend and worsen tissue injury as Shigella does. This phenomenon may be associated with the lower level of virulence factor expression in EIEC when compared to Shigella (Moreno et al. 2009). However, the lower pathogenicity of EIEC could also be related to the host inflammatory response against these bacteria. In addition to studying the expression of VP virulence genes of EIEC and Shigella after internalization into macrophages, we also investigated how macrophages responded to infection by these pathogens. No significant cytokine expression differences were observed in J744 macrophages infected with EIEC or S. flexneri. A comparable elevation in the expression of IL-1, IL-10 and TNF-α was detected in culture supernatants 6 h post-infection with both species. This result is in agreement with the similar expression of ipaB in EIEC and S. flexneri. In conclusion, EIEC demonstrates a delayed killing effect in J774 macrophage cultures when compared to S. flexneri. This in vitro behavior was found to be related to the diminished ipaC expression. Therefore, EIEC's "slow invasion" of colonic epithelial cells in vivo could be related to the hypoexpression of ipaC, leading to a prolonged presence inside infected cells. All together, these data could explain why EIEC takes longer than Shigella species to cause diarrhea. We believe that more experiments with host cells should be conducted to better understand the inflammatory response generated by EIEC. ACKNOWLEDGEMENTS To Dr Patrícia Tobo, for help with real-time PCR analysis. REFERENCES
Copyright © 2010 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc10132f4.jpg] [oc10132f3.jpg] [oc10132f2.jpg] [oc10132t1.jpg] [oc10132t2.jpg] [oc10132f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}