 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
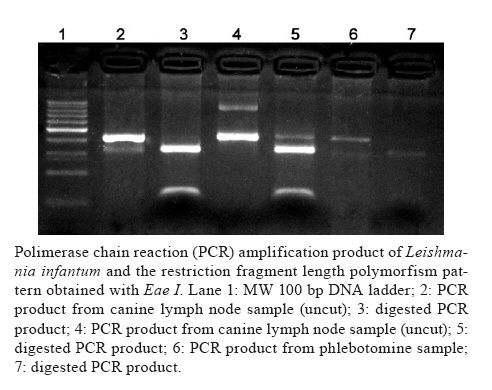
Memórias do Instituto Oswaldo Cruz, Vol. 105, No. 6, 2010, pp. 796-799 ARTICLES Detection of Leishmania infantum in naturally infected Lutzomyia longipalpis (Diptera: Psychodidae: Phlebotominae) and Canis familiaris in Misiones, Argentina: the first report of a PCR-RFLP and sequencing-based confirmation assay Soraya Alejandra AcardiI, +; Domingo Javier LiottaI; María Soledad SantiniII; Carlo Mariano RomagosaIII; Oscar Daniel SalomónII ILaboratorio
de Biología Molecular Aplicada, Facultad de Ciencias Exactas, Químicas
y Naturales, Universidad Nacional de Misiones, Posadas, Misiones, Argentina + Corresponding author: sorayacardil@gmail.com Received 17 February 2010 Financial Support: CeNDIE, CONICET, LaBiMAp Code Number: oc10134 ABSTRACT In this study, a genotypification of Leishmania was performed using polimerase chain reaction-restriction fragment length polymorfism (PCR-RFLP) and sequencing techniques to identify species of Leishmania parasites in phlebotomine sand flies and dogs naturally infected. Between January-February of 2009, CDC light traps were used to collect insect samples from 13 capture sites in the municipality of Posadas, which is located in the province of Misiones of Argentina. Sand flies identified as Lutzomyia longipalpis were grouped into 28 separate pools for molecular biological analysis. Canine samples were taken from lymph node aspirates of two symptomatic stray animals that had been positively diagnosed with canine visceral leishmaniasis. One vector pool of 10 sand flies (1 out of the 28 pools tested) and both of the canine samples tested positively for Leishmania infantum by PCR and RFLP analysis. PCR products were confirmed by sequencing and showed a maximum identity with L. infantum. Given that infection was detected in one out of the 28 pools and that at least one infected insect was infected, it was possible to infer an infection rate at least of 0.47% for Lu. longipalpis among the analyzed samples. These results contribute to incriminate Lu. longipalpis as the vector of L. infantumin the municipality of Posadas, where cases of the disease in humans and dogs have been reported since 2005. Key words:visceral leishmaniasis - Leishmania infantum - Lutzomyia longipalpis - Argentina Visceral leishmaniasis (VL) is the most severe clinical manifestation of leishmaniasis observed worldwide, with an estimated incidence of roughly 500,000 cases per year and a mortality rate of 59,000 deaths per year. The primary parasite, vector and reservoir involved in the transmission cycle in Latin America have been identified as Leishmania infantum, Lutzomyia longipalpis and Canis familiaris (PAHO 2008), respectively. The presence of Lu. longipalpis has been reported in northeastern Argentina (specifically the cities of Candelaria and Corpus, in the province of Misiones) since the 1950s (Salomon et al. 2001). However, it was not until the urban VL outbreaks of 2006 in the state of Mato Grosso do Sul (MS), Brazil, and the city of Asunción, Paraguay, that the first human case of VL was reported in Posadas, a city in the province of Misiones, Argentina. Investigations prompted by this case led to the discovery of Lu. longipalpis in the backyard of the home of the infected individual; this was followed by diagnosis of the disease in several dogs from the same neighborhood. The human case was confirmed by the presence of parasites in bone marrow smears, as well as by culture, serology (rK39 dipstick) and PCR (Leishmania genus); furthermore, parasite isolations were performed in the canine confirmation procedure (Salomon et al. 2008). By December of 2009, 39 human VL cases had been reported in the urban area of Posadas, according to the National System Surveillance of Health database. Apart from immunoserological evidence and amastigote observation in smears, there has been no confirmation of the specific parasite species involved in this outbreak, the southernmost focus of VL known. Therefore, in this study we performed a Leishmania genotypification in phlebotomine sand flies and dogs using polimerase chain reaction-restriction fragment length polymorfism (PCR-RFLP) and sequencing techniques (Aransay et al. 2000, Jorquera et al. 2005, de Pita-Pereira et al. 2005, 2008). The primary goal of this work was to detect and identify the Leishmania species present and to identify putative vectors and reservoirs of these parasites in this focus; such information could lead to a better understanding of the local disease epidemiology and how it relates to the nearby northern VL foci. SUBJECTS, MATERIALS AND METHODS Sand fly and animal samples - Phlebotomine sand flies were captured with CDC minilight traps that were operated overnight. The traps were located at 13 peridomestic environments (27º24'404''S, 55º57'811''W; 27º24'045''S, 55º56'884''W; 27º23'001''S, 55º56'667''W; 27º24'030''S, 55º55'342''W; 27º24'471''S, 55º55,337''W; 27º23'333''S, 55º55'462''W; 27º22'431''S, 55º56'648''W; 27º22'298''S, 55º56'456''W; 27º25'719''S, 55º53'161''W; 27º25'754''S, 55º53'210''W; 27º23'012''S, 55º56'724''W; 27º23'855''S, 55º56'433''W; 27º22'737''S, 55º56'431''W) between 22 January-18 February within the city of Posadas (27°23'S; 55°53'W, 120 m above sea level). Taxonomic identification was performed according to Young and Duncan (1994), with the modification of Andrade Filho et al. (2003); Lu. longipalpis was the only species found. Random selection of sand flies from each trap was collected and stored at -20ºC for molecular analysis. Only traps that contained five or more females were included in the study. The females were assembled in groups (5-12 specimens per group) belonging to the same trap. The percentage of females used for PCR per trap ranged from 52-100% in 12 of the 13 traps, whereas in the remaining trap, 37% of the females were used. Canine samples were collected by a medical veterinarian at the Posadas Municipality Animal Health Institute. Lymph node aspirates used for PCR analysis were collected from two symptomatic stray animals from Posadas that had been positively diagnosed with canine VL; the positive diagnoses were confirmed either by rk39 dipstick or lymph node smear. DNA extraction, PCR-RFLP and cycle sequencing - Sand flies were macerated with a sterile glass micromortar in 1.5 mL tubes containing 500 μL lysis buffer (10 mM tris-HCl, pH 8; 100 mM NaCl; 0.45% Tween 20; 0.25% SDS; 200 μg/sample proteinase K) and incubated at 58ºC for 6 h, followed by a purification step with an equal volume of a mixture of phenol/chloroform/isoamilic alcohol (25:24:1 ratio, pH 8). Overnight DNA precipitation was performed with 100% ethanol and pellets were resuspended in 50 μL DNAse/RNAse-free distilled water according to de Pita-Pereira et al. (2005) with some minor modifications. Lymph node samples were processed with 500 μL lysis buffer (10 mM tris-HCl, pH 8; 100 mM NaCl; 0.45% Tween 20; 0.25% SDS; 200 μg/sample proteinase K) and incubated at 58ºC for 3 h. DNA purification and precipitation steps were performed as previously described for phlebotomine samples. The PCR assay was performed according to the protocol suggested by Marfurt et al. (2003); the described primers Fme (5'-TAT TGGTAT GCG AAA CTT CCG-3') and Rme (5'-ACA GAA ACT GAT ACT TAT ATA GCG-3') that target the mini-exon gene present as tandem repeats in all species of the Leishmania genus were used. The reaction was carried out with 5 μL of each sample in a final volume of 50 μL containing 1X PCR buffer (200 mM tris-HCl, pH 8), 0.1 mM EDTA, 1 mM DTT, 50% glycerol (v/v) (Invitrogen), 1 mM MgCl2 (Invitrogen), 10% DMSO (Sigma), 0.2 mM dNTP Mix, 0.5 μM Fme primer, 0.5 μM Rme primer and 1.4 U Taq polymerase (Invitrogen). Up to 10 μL of the amplification product was analyzed by 2% agarose gel electrophoresis in TBE buffer containing ethidium bromide (0.5 μg/mL). The RFLP assay was performed by digesting 10 μL of the 418 bp PCR product using the Eae I restriction enzyme (5'-Py▼%GGCCPu-3', New England Biolabs). The DNA fragments were resolved by 2.5% agarose gel electrophoresis in TBE buffer containing ethidium bromide (0.5 μg/mL). Sequence confirmation of the 418 bp product was performed by automatic cycle sequencing of both DNA chains using a fluorescent ddNTP terminator system (Macrogen Inc). RESULTS In this study, we performed PCR analysis on a selected sample of female Lu. longipalpis (n = 211) grouped into 28 pools corresponding to 13 capture sites and two canine samples collected from symptomatic dogs. One vector pool of 10 sand flies (Genbank accession GU565337) and both of the symptomatic canine samples (Genbank accessions GU565338 and GU565339) tested positively for L. infantum by PCR and RFLP pattern analysis (Figure). A BLAST search performed against the Genbank database using the PCR product DNA sequences showed a maximum identity with L. infantum. The detection limit of the assay, when performed with L. infantum isolated from local dogs, was determined to be 10 picograms. Assuming that the total DNA content of Leishmania is approximately 100 femtograms per genome (Myler & Stuart 2000, Gontijo & de Carvalho 2003), the positive pool analyzed in this study contained nearly 100 parasites. Although the sand fly sample is not representative of the total population of sand flies in Posadas, we found a 3.5% rate of L. infantum infection among the pools of macerated Lu. longipalpis females; when taking into account the number of individual specimens tested, we determined an infection rate of at least 0.47% (1 out of 28 pools, with at least 1 infected insect in the positive pool). Previous studies using various PCR-based techniques to determine region-specific Lu. longipalpis infection rates in MS determined a rate of 0.25% in Bonito (Savani et al. 2009), whereas in the older settled foci in Campo Grande and Corumbá, it reached 1.9% for Leishmania spp (Silva et al. 2008) and 1.5% for the Lu. longipalpis sibling species (and proven vector of L. infantum) Lutzomyia cruzi (de Pita-Pereira et al. 2008), respectively. Aside from differences in the sampling strategies and techniques used, the differences could be related to the eco-epidemiology of each urban VL foci. The spatial distribution of Lu. longipalpis in Posadas resembled a clustered pattern (Fernández et al. 2010); the infection and transmission rates could be higher in these "patches" of vector abundance. One of the five vector incrimination criteria suggested by Killick-Kendrick (1990) is the simultaneous identification of Leishmania genus in patients and sand flies in a given area. This requires parasite visualization in both the vector digestive tract and mammalian samples, which would typically be followed by culture isolation and other time-consuming laboratory techniques (Aransay et al. 2000, Santamaría et al. 2005). Although these techniques are considered the gold standards, in the last two decades the development of PCR and other molecular biology assays have made testing in less-advanced laboratories practical and have enabled the use of highly sensitive diagnostic tools in areas where such diseases strongly impact the population. In particular, the usefulness of these techniques with respect to leishmaniasis lies in their ability to quickly identify Leishmania species, allowing for better surveillance of the vector directly involved in the transmission cycle (Cruz et al. 2006, Moreira et al. 2007, de Pita-Pereira et al. 2008). DISCUSSION In conclusion, though parasite DNA detection is an indirect method of isolation (Alvar Ezquerra 2001) that does not distinguish between the presence and viability of parasites, sequence-based species confirmations were consistent with previous immunoserological results using samples from humans and dogs in the same geographic area (Salomon et al. 2008). Therefore, for the first time in Argentina, the local genotypification of L. infantum by RFLP-PCR, together with the direct observation of amastigote forms in lymph node smears, implicated dogs as reservoirs and Lu. longipalpis as potential vectors in this urban focus. This scenario is similar to those described in recently developed foci that formed during the dispersion of VL to the southern cities of South America through Brazil (Silva et al. 2008) and Paraguay (Cousiño 2006) prior to reaching this southernmost focus in Argentina. The similarities between these foci validate the importance of designing regional control strategies. Likewise, the clustered distribution of sand flies observed in Posadas (Fernández et al. 2010) and the infection rates presented in this report highlight the need for research on parasite circulation in these spatially heterogeneous urban scenarios to improve the current control approaches. Furthermore, the proper identification of parasite species through highly sensitive and specific alternative techniques such as RFLP-PCR is of epidemiological and clinical importance. This is particularly true in areas where Leishmania amazonensis could be present, such as Argentina (Frank et al. 2003), as well as MS (Lima Junior et al. 2009), where it could be infecting Lu. longipalpis (Savani et al. 2009), a species that is also capable of transmitting VL (Almeida et al. 1996, Aleixo et al. 2006, Tolezano et al. 2007). ACKNOWLEDGEMENTS To Dr Lilian Tartaglino (Secretaria Calidad de Vida) and the technicians of the Entomology Laboratory of the Municipality of Posadas, for providing local support during the sampling. REFERENCES
Copyright © 2010 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc10134f1.jpg] |
| |||||||||

{kind=link}