 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
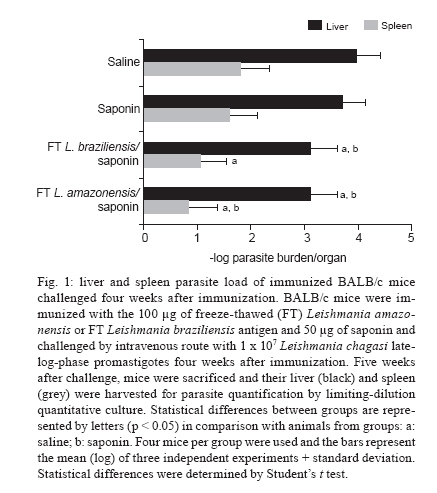
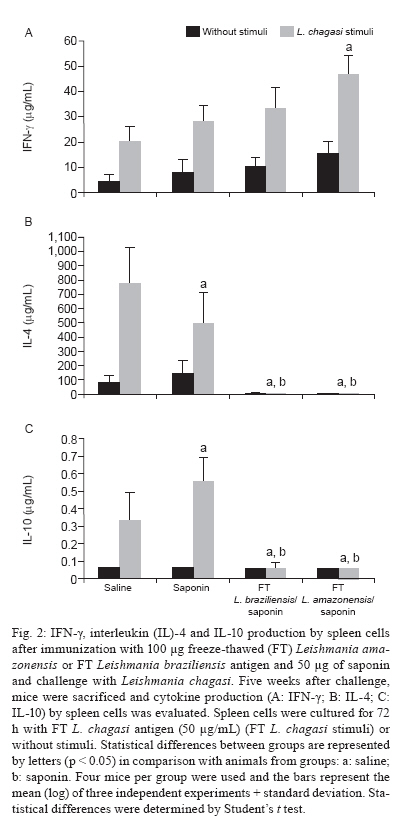
Memórias do Instituto Oswaldo Cruz, Vol. 105, No. 6, 2010, pp. 823-828 ARTICLES Antigenic extracts of Leishmania braziliensis and Leishmania amazonensis associated with saponin partially protects BALB/c mice against Leishmania chagasi infection by suppressing IL-10 and IL-4 production Rafaella FQ GrenfellI, II, +; Eduardo A Marques-da-SilvaII; Miriam C Souza-TestasiccaII; Eduardo AF CoelhoIII, IV, V; Ana Paula FernandesIII; Luís Carlos C AfonsoII; Simone A RezendeII, VI ILaboratório
de Esquistossomose, Instituto de Pesquisas René Rachou-Fiocruz, Av. Augusto
de Lima 1715, 30.190-002 Belo
Horizonte, MG, Brasil + Corresponding author: rafaella@cpqrr.fiocruz.br Received 23 March 2010 Code Number: oc10138 ABSTRACT This study evaluated two vaccine candidates for their effectiveness in protecting BALB/c mice against Leishmania chagasi infection. These immunogenic preparations were composed of Leishmania amazonensis or Leishmania braziliensis antigenic extracts in association with saponin adjuvant. Mice were given three subcutaneous doses of one of these vaccine candidates weekly for three weeks and four weeks later challenged with promastigotes of L. chagasi by intravenous injection. We observed that both vaccine candidates induced a significant reduction in the parasite load of the liver, while the L. amazonensis antigenic extract also stimulated a reduction in spleen parasite load. This protection was associated with a suppression of both interleukin (IL)-10 and IL-4 cytokines by spleen cells in response to L. chagasi antigen. No change was detected in the production of IFN-γ. Our data show that these immunogenic preparations reduce the type 2 immune response leading to the control of parasite replication. Key words: vaccine - Leishmania chagasi - Leishmania braziliensis - Leishmania amazonensis - IL-10 - IL-4 Leishmaniasis is a spectrum of diseases caused by infection with different Leishmania species that range from self-limiting cutaneous leishmaniasis to visceral leishmaniasis (VL). VL, also known as kala-azar, is fatal if not successfully treated. Human infection with Leishmania chagasi/Leishmania infantum, the protozoan causing South American VL, ranges from sub-clinical infection to progressive fatal disease (Wilson 1993). Sub-clinical infection results in the development of a cellular immune response that often results in long-term protective immunity against re-infection (Pearson & Sousa 1996), suggesting that a vaccine against leishmaniasis is feasible. Since treatment of VL is difficult and no acceptable vaccine exists against human infection, the development of an effective vaccine would be an important achievement. An ideal vaccine would be one that could offer cross-protection against diverse Leishmania species (Campbell et al. 2003). The involvement of T helper 1 (Th1) and T helper 2 (Th2) subsets with either protection or disease exacerbation has been demonstrated in murine cutaneous leishmaniasis (Heinzel et al. 1989). A similar pattern of Th cell subsets has been shown in some studies for VL (Rhee et al. 2002), mainly because interleukin-4 (IL-4) can regulate macrophage function (Hamilton et al. 1999). Unexpectedly, some studies in animal models have proven that protection in VL is associated with the production of both type 1 and type 2 cytokines (Ghosh et al. 2001, Ramiro et al. 2003, Vilela et al. 2007). In this study, we evaluated the potential of two freeze-thawed (FT) Leishmania antigenic extracts for protection against L. chagasi infection. Cross-species protection has been supported in many studies on leishmaniasis (Bebars et al. 2000, Misra et al. 2001, Tonui & Titus 2007). Therefore, we decided to investigate whether subcutaneous immunization with extracts of Leishmania amazonensis or Leishmania braziliensis could protect BALB/c mice against L. chagasi infection. These two antigenic extracts and a purified saponin fraction from the bark extracts of Quillaja saponaria, which has been considered a promising adjuvant in numerous prophylactic and therapeutic vaccines, were used in association (Kensil 1996). It is worth noting that the mechanism by which saponin potentiates an immune response remains unclear. Hypotheses have been raised about whether saponin, through lectin-mediated cell membrane interactions, could facilitate the uptake of the antigen into antigen-presenting cells, leading to specific cytokine profiles that enhance T and/or B-cell responses (Kensil 1996, Marciani 2003, Adams et al. 2010). We decided to evaluate these two species because a tested vaccine composed of L. amazonensis along with Bacillus Calmette-Guérin has been effective in the treatment of cutaneous leishmaniasis patients in Venezuela (Convit et al. 2003). Further, a recent vaccine composed of L. braziliensis, sand fly gland extract and saponin was shown to be immunogenic in dogs (Giunchetti et al. 2007, 2008). These species are associated not only with localized cutaneous leishmaniasis, but also with mucocutaneous leishmaniasis and anergic diffuse cutaneous leishmaniasis in Brazil. This study aimed at evaluating whether these two vaccine candidates, composed of antigens obtained from species responsible for cutaneous and mucocutaneous leishmaniasis, could protect against murine VL caused by L. chagasi. MATERIALS AND METHODS Leishmania parasites and antigens - The strain of L. chagasi used in this study (MHOM/BR/1974/M2682) was kindly provided by Dr Maria Norma de Melo, from the Parasitology Department, Federal University of Minas Gerais (UFMG), Brazil. Promastigotes were grown in Dulbecco's Modified Eagle Medium (DMEM; pH 6.8) supplemented with 20% heat-inactivated foetal bovine serum (FBS), 2 mM L-glutamine, 25 mM HEPES, 50 μM 2-mercaptoethanol and 20 μg/mL garamicin [DMEM 20% phosphate buffered saline (PBS)] at 25ºC. Infectivity was maintained by serial passage through mice. L. amazonensis strain PH8 (IFLA/BR/67/PH8) and L. braziliensis strain M2903 (MHOM/BR/75/M2903) were grown in Grace's Medium supplemented with 20% FBS, 2 mM L-glutamine and 20 μg/mL garamicin. Promastigotes of L. chagasi, L. amazonensis or L. braziliensis were harvested from late-log-phase cultures by centrifugation, washed three times with PBS and disrupted by three rounds of freezing and thawing. The protein contents were estimated (Lowry et al. 1951) and the antigen was frozen at -70ºC prior to use. L. amazonensis and L. braziliensis antigens were used to immunize mice. Mice, immunizations and the quantification of parasite load - Female BALB/c mice (4-6 weeks old) were obtained from Bioterism Center, UFMG, and were maintained at the Central Biotherium, Federal University of Ouro Preto (UFOP). Four BALB/c mice per group, in three independent experiments, were injected weekly with three subcutaneous doses of 100 μg of L. amazonensis or L. braziliensis antigenic extracts together with 50 μg of saponin as an adjuvant. Control mice were inoculated with PBS or 50 μg of saponin alone. Four weeks later, mice were challenged with 1 x 107 promastigotes of L. chagasi given intravenously through the lateral tail vein. Five post challenge mice were sacrificed and the spleen and liver parasite loads were determined by quantitative limiting-dilution culture. Quantitative limiting-dilution culture was performed as previously described with some modifications (Marques-da-Silva et al. 2005). Briefly, spleen and liver were harvested and weighed. One fragment of each organ was obtained and weighed separately for parasite quantification. This fragment was homogenized in a tissue grinder, resuspended in 500 μL of DMEM containing 20% FBS and plated onto 48-well flat-bottom microtiter plates. Five-fold serial dilutions were performed and, after two weeks of incubation at 25ºC, plates were microscopically scored for parasite growth. The number of parasites was determined from the reciprocal of the highest dilution at which promastigotes could be detected and is expressed as parasites per organ. Determination of vaccine-induced cytokine production - Single-cell suspensions of spleen were obtained by homogenization in a tissue grinder. The erythrocytes were lysed with ammonium chloride lysis buffer and the cells were washed. Cells were then cultured in DMEM (pH 7.2) supplemented with 10% heat-inactivated FBS, 2 mM L-glutamine, 25 mM HEPES, 50 μM 2-mercaptoethanol and 20 μg/mL garamicin (DMEM 10% FBS) at 5 x 106 cells/mL in 48-well flat-bottom plates without stimuli (ws) or stimulated with 50 μg of L. chagasiAg/mL of culture for 72 h. The production of IFN-γ and IL-4 was determined by the presence of these cytokines in cell culture supernatant, as measured by enzyme linked immunosorbent assay (ELISA) using specific purified monoclonal antibodies (Afonso & Scott 1993). The production of IL-10 was assayed using a commercial ELISA kit, according to the manufacturer's instructions (Duo Set®, R&D Systems). Statistical analysis - Parasite burden data were logarithmically transformed to determine the homogeneity of variance. All data were analyzed using the Kolmogorov-Smirnov normality test. Data with a normal distribution were analyzed by the Student's t test. Ethics - This study was approved by the Ethical Committee of the UFOP. RESULTS Parasite load in liver and spleen - In order to determine if L. amazonensis or L. braziliensis antigenic extracts were able to protect BALB/c mice from L. chagasi infection, we evaluated the parasite load within the liver and spleen by limiting dilution analyses. Mice that were immunized with FT L. amazonensis antigen showed significant reduction in parasite load in the liver and spleen (p < 0.05), as shown in Fig. 1. The L. braziliensis antigen was also able to reduce the parasite load in the liver but did not significantly decrease the parasite load in the spleen. Determination of IFN-γ, IL-4 and IL-10 production by spleen cells after vaccination and challenge - In order to determine if these vaccine candidates could influence the immune response to L. chagasi, spleen cells from immunized animals were obtained and incubated with 50 μg/mL of FT L. chagasi antigen or cultured ws. Fig. 2A shows that although no change in IFN-γ production was noted following immunization, spleen cells from animals treated with either vaccine candidate exhibited significantly reduced production of IL-4 (Fig. 2B) and IL-10 (Fig. 2C) as compared to control spleen cells (p < 0.05). DISCUSSION In recent years, several efforts have been made to obtain a safe and efficient vaccine against leishmaniasis. Vaccination with live, attenuated parasites has been attempted (Streit et al. 2001, Nylén et al. 2006), although there are several ethical considerations regarding these vaccines. As such, attention has shifted to the use of recombinant or synthetic antigens, purified fractions or whole antigenic extracts of parasites as an alternative to live parasites. Whole antigenic extract obtained from parasites is a reasonable alternative considering its immunogenicity, cost and safety. Furthermore, any human vaccine will probably require several different antigens and adjuvant to guarantee a satisfactory response by a majority of the population, given its heterogeneity (Handman 2001). A vaccine against cutaneous leishmaniasis was developed by Mayrink et al. (1979). It was prepared from whole parasite antigens obtained from five stocks of parasites isolated from patients with different forms of leishmaniasis. Subsequently, the same group developed a second vaccine based only on the PH8 strain of L. amazonensis. This vaccine has been used in the prevention of disease, as well as serving as an immunotherapeutic agent, thereby demonstrating that administration of the vaccine in association with antimonium salts could be therapeutic. Indeed, when compared with conventional therapy, L. amazonensis vaccine treatment reduced the time necessary for lesions in patients with cutaneous leishmaniasis to completely heal (Toledo et al. 2001). Furthermore, Mayrink et al. (2006) have shown that the association of a vaccine antigen with antimonium salts reduces both the total salt volume and the treatment length, thereby reducing the side effects otherwise associated with the use of antimonium salts. During this study, we evaluated whether antigens from L. amazonensis and L. braziliensis could provide heterologous protection against L. chagasi infection in BALB/c mice. These antigens were used in conjunction with saponin, an adjuvant that has been used in studies involving VL or cutaneous leishmaniasis in mice and dogs (Santos et al. 2002, Nico et al. 2007, Fernandes et al. 2008, Borja-Cabrera et al. 2010). We found that both vaccine candidates were able to reduce parasite load in the liver, but that only the L. amazonensis immunogenic extract reduced the parasite load of the spleen. These data suggest that different mechanisms are utilized to afford protection by these freeze-thawed vaccines. In mice, Vilela et al. (2007) have shown that a vaccine composed of L. amazonensis (PH8 strain) and Corynebacterium parvum is able to protect against L. chagasi infection. Although this vaccine used antigens derived from L. amazonensis, a different pattern of cytokine expression was observed, since in this case protection was associated with an increase in both type 1 and type 2 cytokines. Studies with L. braziliensis have shown that a vaccine composed of FT antigen and saponin may prevent L. chagasi infection in dogs (Giunchetti et al. 2007, 2008). In order to assess the immune response induced by both vaccine candidates, we evaluated the production of cytokines by spleen cells. Although we detected a difference in the pattern of protection between the two organs tested, none of the immunogenic preparations led to increased IFN-γ production. In contrast both antigenic preparations resulted in suppressed IL-10 and IL-4 production by spleen cells. That the suppression of cytokines may reduce parasite load has previously been noted in spleen following the immunization of mice with Leishmania major antigens and intravenous challenge with Leishmania donovani. These mice had reduced IL-4 and IL-10 cytokine levels together with an increase in IFN-γ production. This particular cytokine pattern was not observed when the same immunization was performed together with a L. braziliensis infection. Why this occurred is a matter of debate. However, certain possibilities seem plausible: first, there may be something inherently different about L. braziliensis as compared to L. donovani and their interactions with the immune system that elicit distinct levels of protection. For instance, each of these parasites may differ in their ability to induce effector vs. regulatory T cells (Tonui & Titus 2007). Saponin adjuvant could influence the generation of an immune response at several levels: these may include the mobilization of appropriate antigen-presenting cells to the injected site, enhancing efficient antigen processing and presentation, influencing the cytokine response, including IFN-γ and the co-stimulatory signals necessary for an optimal immune response and increasing the recruitment of effector immune cells to the inflammatory areas (Caro et al. 2003, Buendía et al. 2007). The capacity of this adjuvant to elicit a strong CD8 T cell response has also been reported (Newman et al. 1992). These studies emphasize the immunostimulatory capability of saponin that may have led to the improved level of protection found in our study, although saponin alone did not lead to enhanced IFN-γ production by spleen cells. The effect of our vaccines on IL-10 production has been observed in other studies. Uzonna et al. (2004) had shown that vaccination with L. major mutants was associated with a decrease in IL-10 and IL-4 production. Similarly, Bhaumik et al. (2009) showed that a vaccine candidate composed of soluble antigen from attenuated L. donovani promastigotes was able to provide complete protection against experimental VL and that this protection was associated with a decrease in the production of IL-10. Gomes et al. (2007) showed that, after intranasal immunization with a plasmid expressing the Leishmania analogue of the receptors of activated C kinase, BALB/c mice developed lower parasite burdens and had a decrease in IL-10 production. In another study, it was shown that BALB/c mice immunized with a plasmid encoding the A2 gene were protected against experimental challenge with L. amazonensis or L. donovani and that this protection was associated with a reduced level of IL-10 production (Zanin et al. 2007). Finally, it is already known that human VL is associated with high levels of IL-10. As such, suppression of this cytokine might be important for disease because it is involved in the suppression of macrophage activity (Nylén & Sacks 2007). The role of IL-4 in VL is not as well understood. Some studies have shown that IL-4 is important for granuloma maturation and anti-leishmanial activity in the murine model of L. donovani infection (Kemp et al. 1996). In contrast, another study has shown that protection from leishmaniasis is associated with a reduction of type 2 cytokines, including IL-4 (Alves et al. 2009). The current study shows that immunogenic preparations composed of L. amazonensis or L. braziliensis partially protect BALB/c mice from intravenous challenge with L. chagasi promastigotes and that this protection is associated with a reduction in the level of IL-10 and IL-4 expression. The role played by saponin in a model whereby regulatory cytokines are suppressed is not known and should be the subject of additional investigation. Furthermore, it shows that cross-protection between Leishmania species presents a major practical implication because vaccination procedures based on the use of a vaccine from one species will likely protect against different Leishmania species. ACKNOWLEDGEMENTS To Dr Alexandre Barbosa Reis, for his assistance. REFERENCES
Copyright © 2010 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc10138f1.jpg] [oc10138f2.jpg] |
| |||||||||

{kind=link}
{kind=link}